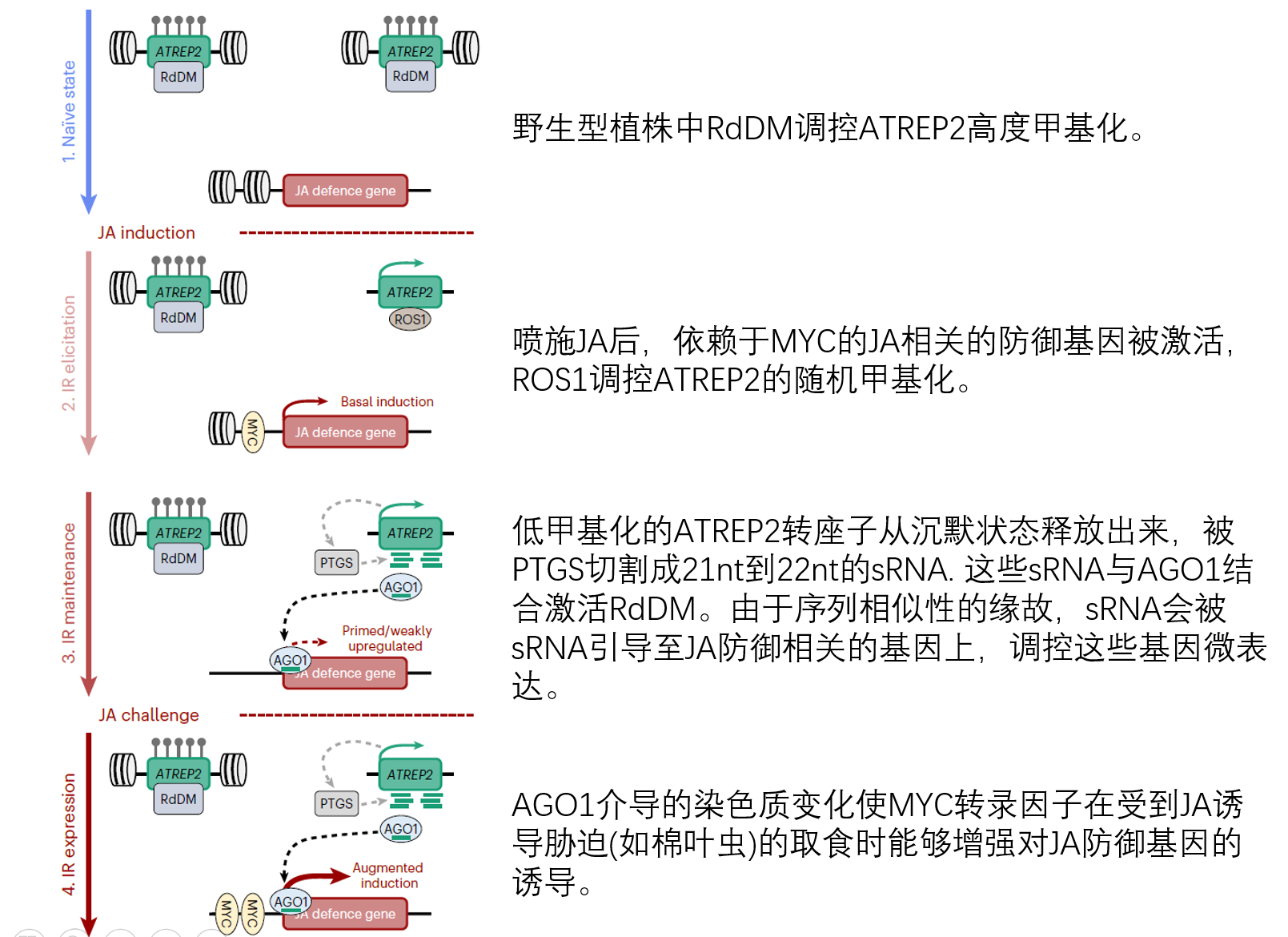
依赖茉莉酸的长久免疫记忆需要DNA甲基化的参与
文献信息
研究背景
- 水杨酸(SA)参与的防御反应主要对(半)活体营养型病原菌发挥作用,茉莉酸(JA)参与的防御反应则主要是对死体营养型病原菌和草食动物发挥作用。
- 植物中胞嘧啶的甲基化通常叫做DNA甲基化,会发生在三种序列上:CG、CHG和CHH(H是除G 以外的其他碱基)。DN甲基化主要作用于转座元件以减少其对基因组的潜在影响。
- RNA靶向的DNA甲基化(RdDM)和DNA去甲基化酶ROS1的拮抗作用共同控制TE富集区DNA甲基化的发生。
- RdDM不仅和翻译后的基因沉默(PTSG)有关,还和小干扰RNA(siRNA)有关。这些siRNA来自依赖RNA聚合酶Ⅱ的转录本,将初始的DNA甲基化位点靶向未甲基化的位点。
- RNA聚合酶Ⅳ也是RdDM所需要的。
- 近些年的研究表明RdDM和ROS1参与植物对生物胁迫的响应。
- JA对植物免疫的短期影响的研究很多,但是其对植物的长期影响及其机制还不清楚。
研究结果
JA的长期影响
JA能够诱导植物对植食性动物的长期抗性,但是对死体营养型病原菌及半活体营养型病原菌却是长期的敏感性。简单来说就是JA诱导产生的长期抗性只对植食性动物有用,对病原菌的作用是负向的。
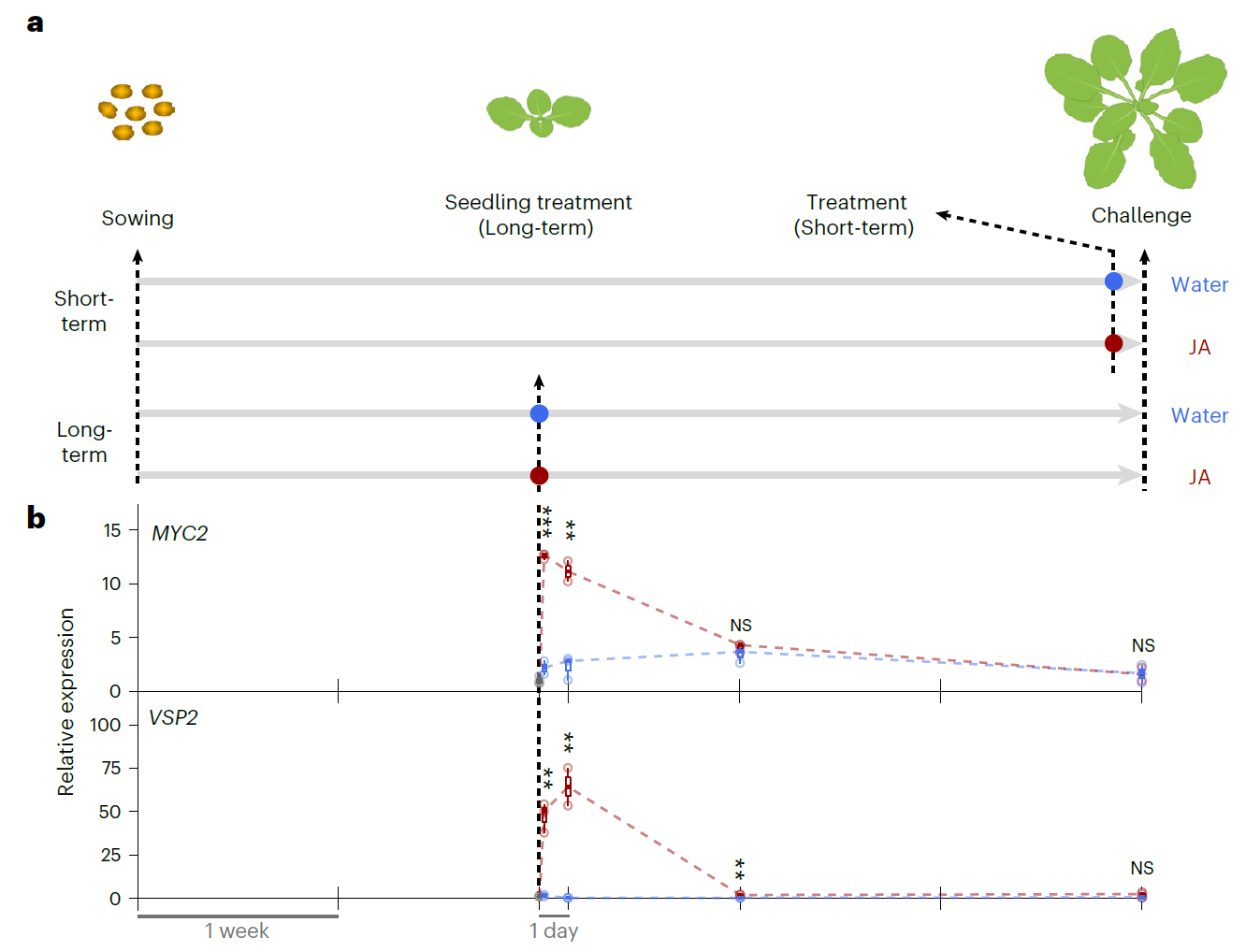
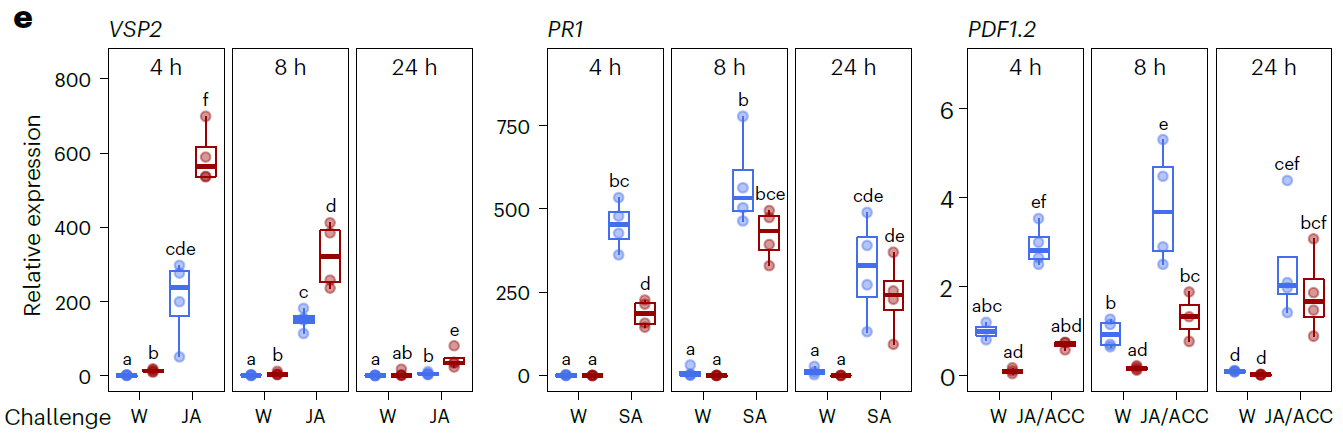
试验设计是比较简单的(下图)。用1mM的茉莉酸处理2周龄的拟南芥幼苗,对照是水,这个处理用作观测诱导产生的长期抗性。一直生长到第五周,这时候开始处理短期诱导抗性分组,同样也是1mM的茉莉酸进行处理,对照同样是水。检测的指标是基因表达量。所选的基因MYC2和VSP2都是和茉莉酸紧密相关的关键基因。采样的时间点是短期处理的在喷施茉莉酸后4小时第一次采样,24小时第二次采样。长期观测处理组则是在处理后1周和3周进行采样。从基因的表达情况可以看出,茉莉酸处理后4小时和24小时,与对照相比,两个基因的表达量都显著的增强了。MYC2在处理后1周和3周其表达量没有法神显著的变化,VSP2在处理后1周其表达量与对照相比显著的增强了(图中看起来是和基线持平的),在处理后3周其表达量与对照相比其表达量没有发生显著变化。
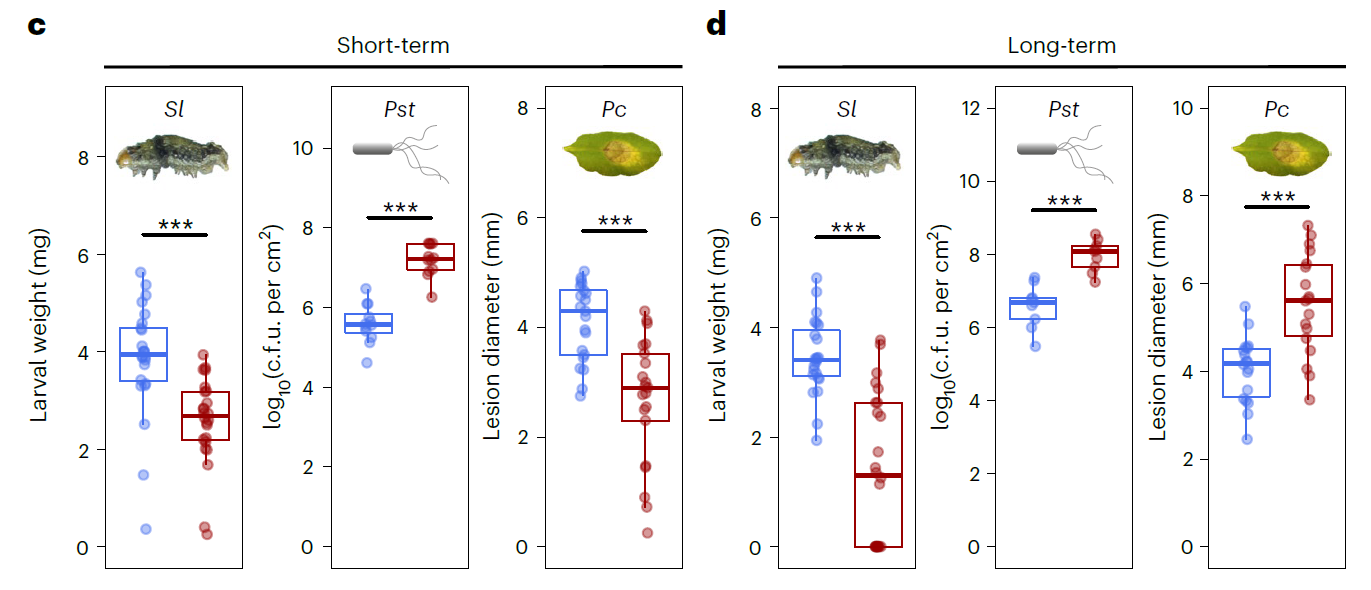
上述结果似乎表明JA诱导的长期免疫没有用,至少在基因表达层面是没有差异的。那么在表型上有没有差别呢?为了验证JA诱导的短/长期免疫的表型差异,设计了如下试验(下图)。检测的对象有植食性昆虫棉叶虫(Spodoptera littoralis, Sl)、半活体营养型病原菌(Pseudomonas syringae pv. tomato DC3000,Pst)和死体营养型病原菌(Plectosphaerella cucumerina,Pc)。简单描述就是第五周的时候,也就是完成长期和短期处理后,分别用Sl、Pst和Pc处理拟南芥叶片,然后测量棉叶虫Sl的体重、Pst在叶片上的菌群数量以及Pc病斑直径。结果发现,不管是短期诱导还是长期诱导,棉叶虫的体重都减轻了,半活体营养型病原菌Pst的菌群数量都增加了。有趣的是,短期诱导时,死体营养型病原菌Pc的菌落直径显著变小;茉莉酸长期诱导后,死体营养型病原菌Pc的菌落直径却增加了。JA诱导的免疫不管是长期还是短期,其作用对植食性昆虫Sl的抑制作用一直存在;JA诱导的短期免疫对死体营养型病原菌Pc是能够正常发挥作用的,但是诱导产生的长期免疫对死体营养型病原菌Pc却有着相反的作用。JA信号参与的免疫过程会抑制依赖于SA的免疫作用,进而影响对(半)活体营养型病原菌的免疫反应,所以不管是JA的短期诱导还是长期诱导,其诱导的免疫对半活体营养型病原菌Pst的抗性均降低了。JA短期诱导的免疫和长期诱导的免疫对死体营养型病原菌Pc有着相反的免疫反应,这表明在拟南芥中,JA诱导的短期免疫和长期免疫有着本质差别。
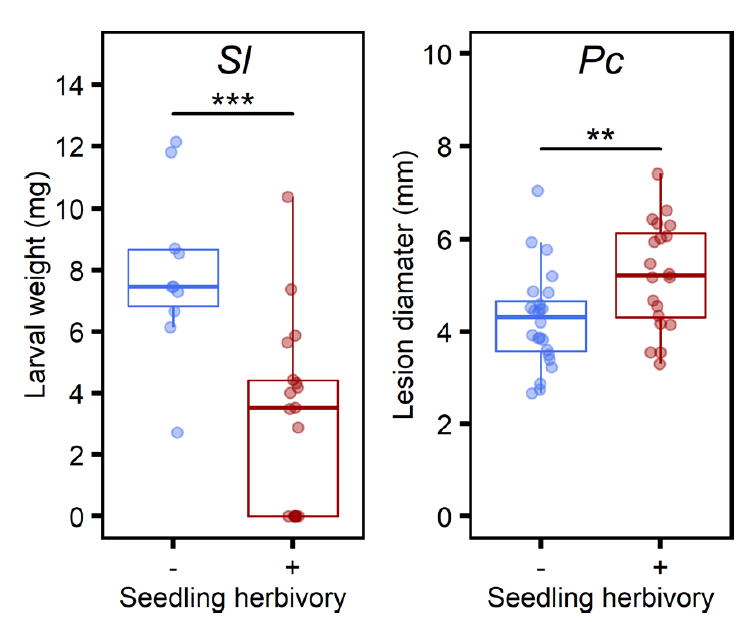
考虑到上述试验的处理都是外源喷施JA完成的,为了验证真实情况下JA诱导的免疫差异,便用棉叶虫Sl取食2周龄的拟南芥幼苗,三周后再用棉叶虫Sl取食拟南芥叶片、死体营养型病原菌Pc侵染拟南芥叶片。结果和外源喷施JA的结果一致:JA诱导的长期免疫对棉叶虫Sl有抑制作用,对死体营养型病原菌Pc的侵染具有促进作用(下图)。
JA诱导的免疫对其他免疫途径的影响
从第一个基因表达的结果可以看出的是,JA长期诱导后的第三周,MYC2和VSP2的表达量与对照相比没有差异,基本与基线持平。对此,作者猜测JA诱导的长期免疫之所以能够对棉叶虫Sl产生抗性,是由于JA诱导的长期免疫激发了依赖于JA途径的相关基因的转录表达。为此,选择依赖于MYC的抗虫相关的酸性磷酸酶基因VSP2进行检测。处理很简单,以水为对照利用JA处理拟南芥幼苗,然后在不同时间点进行新的处理(喷水或者是JA)。先用JA诱导过的拟南芥幼苗再遇到JA的时候,VSP2的表达量显著的增强。这就表明JA在拟南芥幼苗时期诱导产生的免疫能够长时间保持抗虫性。但是有趣的是,结果JA处理诱导后,在受到SA刺激时,SA免疫途径相关基因PR1的表达量显著的降低了;依赖于JA/ET途径的抗真菌相关基因PDF1.2的表达量也显著的降低了。这个结果表明JA诱导的免疫会负向调控SA途径基因PR1及JA/ET途径抗真菌基因PDF1.2的表达。
苗期JA处理对转录组的长期影响
JA诱导的长期免疫会增强植物依赖于JA信号对植食性昆虫的抗性;但是JA诱导的长期免疫会抑制依赖于SA和ET的信号通路,进而降低拟南芥对半活体型病原菌及死体型病原菌的抗性。
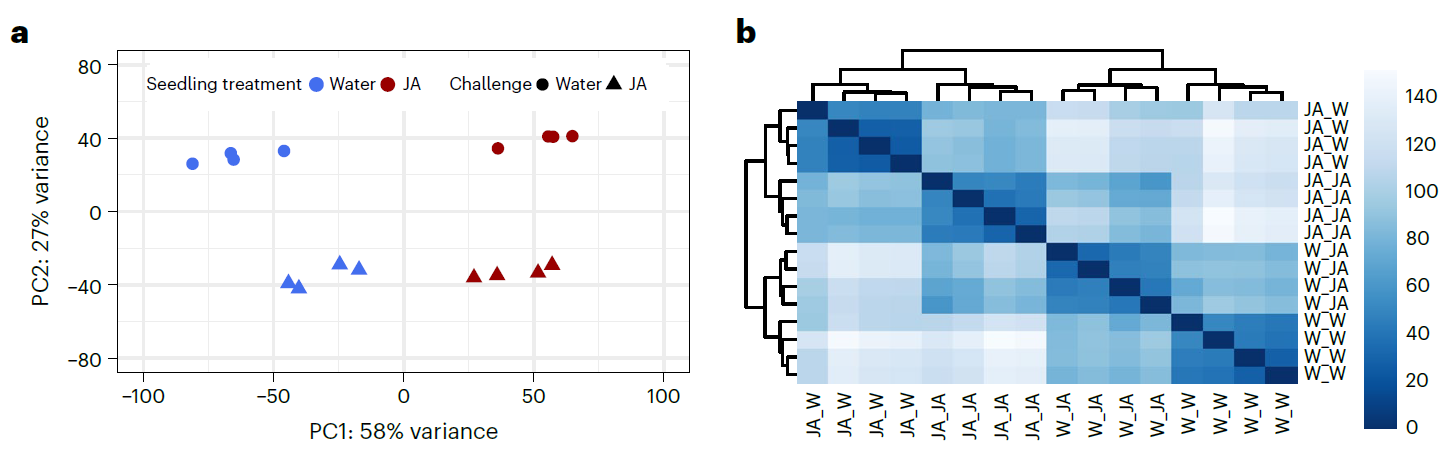
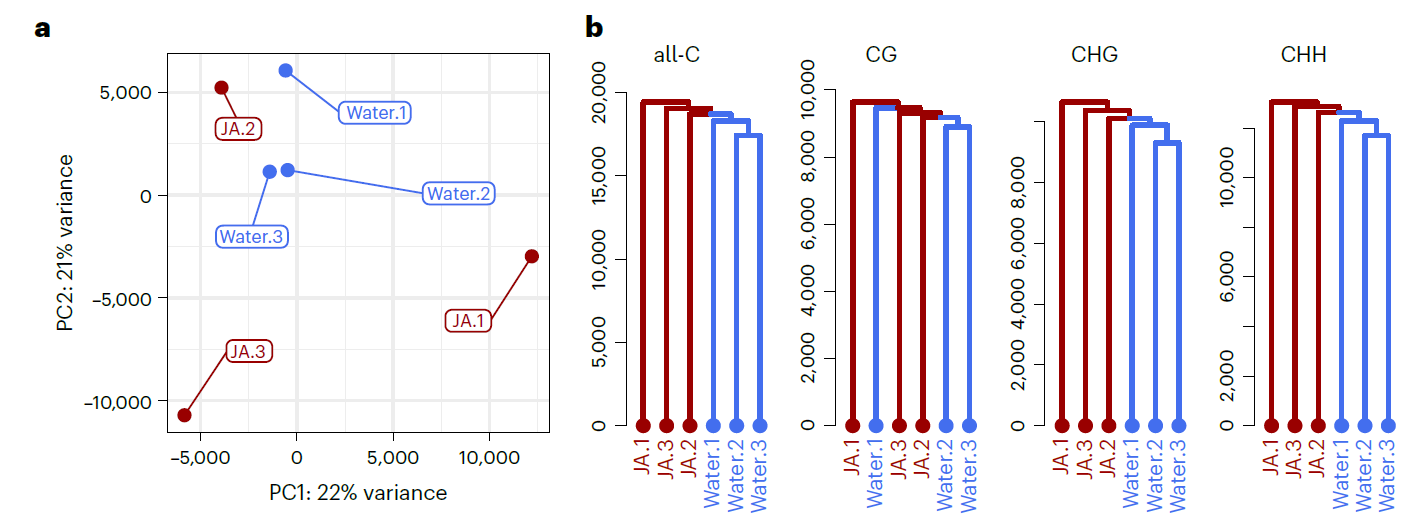
为了验证JA诱导作用对转录组的长期影响,设计了和上面一样的试验:以水为对照,先用JA处理两周龄的拟南芥幼苗;到第五周时在分别用水和JA交叉喷施处理后的拟南芥幼苗,处理4小时后采样进行RNA-Seq. 从PCA和聚类的结果来看,(预)处理间有着很明显的差异。预处理和后续处理之间都有着很明显的差别。从PCA结果可以看出,JA预处理对转录组的影响比后续JA处理的影响更大。
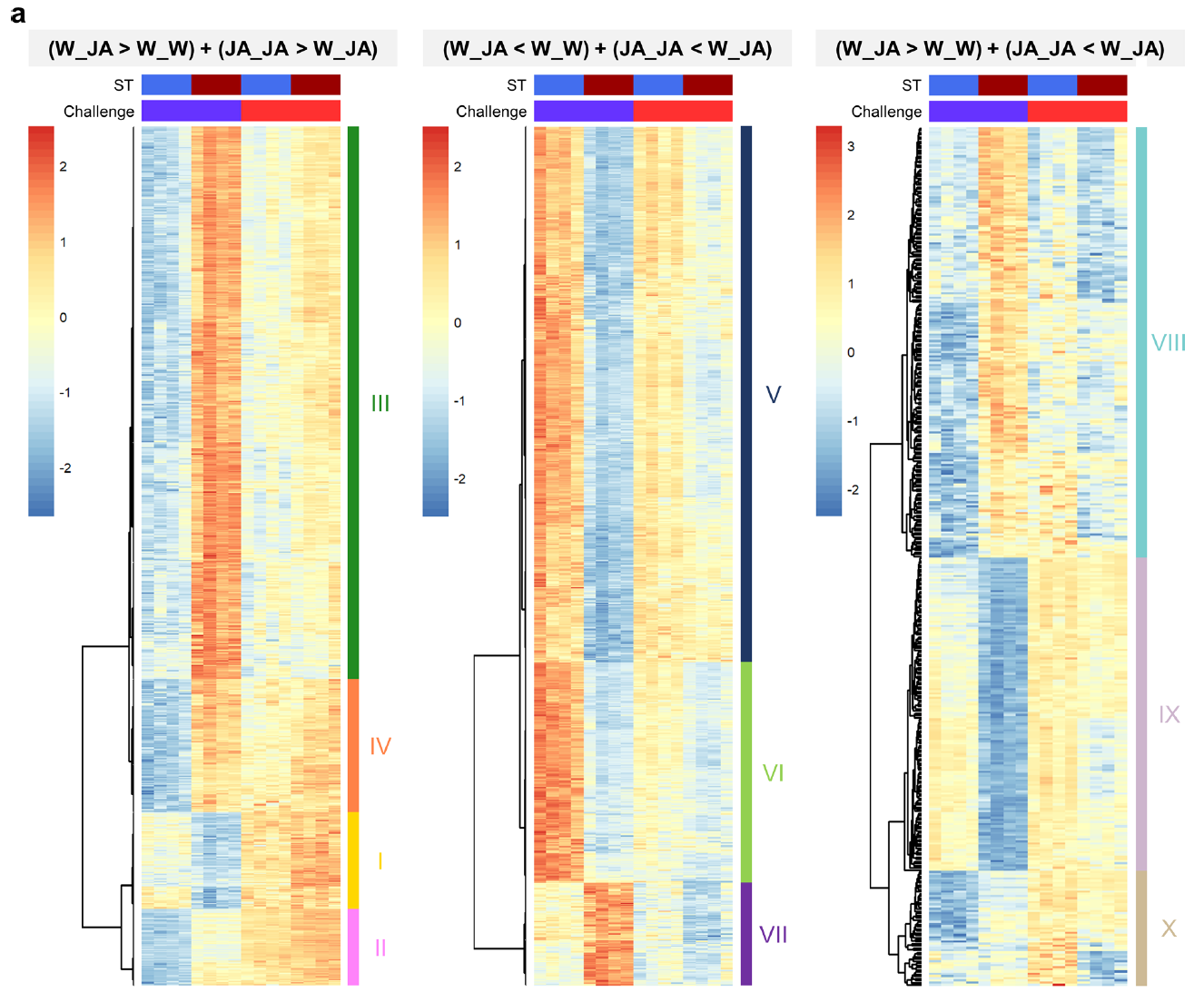
考虑到JA预处理改变了植株对死体营养型病原菌Pc的抗性/敏感性,作者猜测JA预处理改变了JA再次处理时的转录重编程。为此,从转录组数据中筛选出在JA预处理和二次处理之间存在显著差异的2409个基因。针对这2409个基因,进行进一步的筛选:
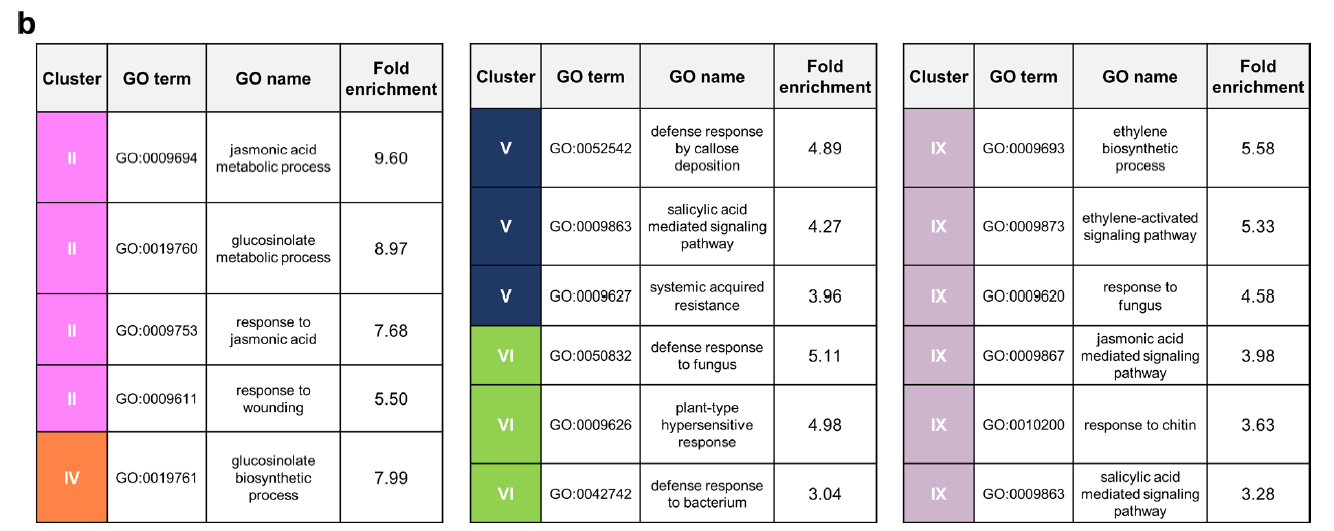
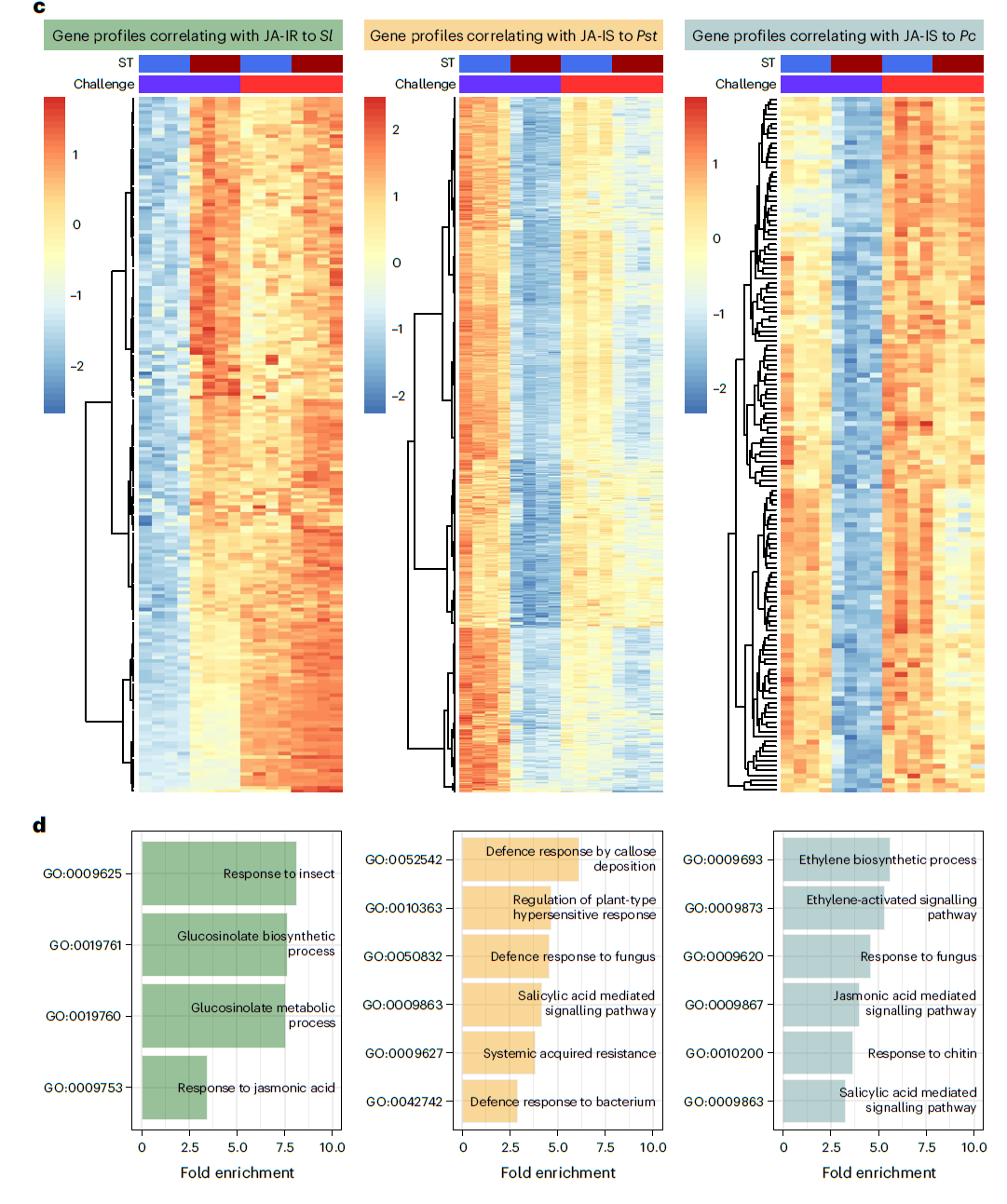
- JA诱导的长期免疫对棉叶虫Sl具有抗性的基因:JA二次处理后其表达量显著上调的基因(W_JA>W_W)和两次JA处理后表达量高于先用水后用JA处理的基因(JA_JA>W_JA)(下图左)(第三列表达量大于第一列,第四列表达量大于第三列)。一共筛选到832个基因,将这些基因进行聚类,一共可以聚为4。在这4类基因中,Ⅱ和Ⅳ类基因表现出明显的JA长期诱导免疫趋势。VSP2也在其中,将这些基因进行GO富集分析,显著富集的GO term包括茉莉酸相关通路、损伤相关通路及芥子油苷生物合成过程等。这些GO term均与植食性动物取食引起的免疫紧密相关。
- JA诱导后对半活体营养型病原菌Pst抗性降低的基因:第二次JA处理后显著下调的基因(W_JA < W_W)(下图中)(第三列表达量小于第一列)和两次JA处理后与单独第二次JA处理相比显著下调的基因(JA_JA<W_JA)(第三列表达量大于第四列)。一共筛选到904个基因,这些基因可以聚类成三类。将这904个基因进行GO富集发分析,发现这些基因大多与水杨酸、植物类过敏性坏死反应及对细菌性病原菌的响应等GO term相关。
- JA长期诱导后对死体营养型病原菌Pc抗性降低的基因:第二次JA处理后表达显著下调的基因(下图右)(W_JA>W_W)(第三列表达量大于第一列)和两次JA处理后表达显著下调的基因(JA_JA<W_JA)(第四列表达量小于第三列表达量)。一共筛选到395个基因,其中144个基因的表达模式符合要求。将这144个基因进行GO富集分析发现这些基因与水杨酸信号、茉莉酸及乙烯等信号通路相关,还有一个GO term与植物对真菌的响应显著相关。
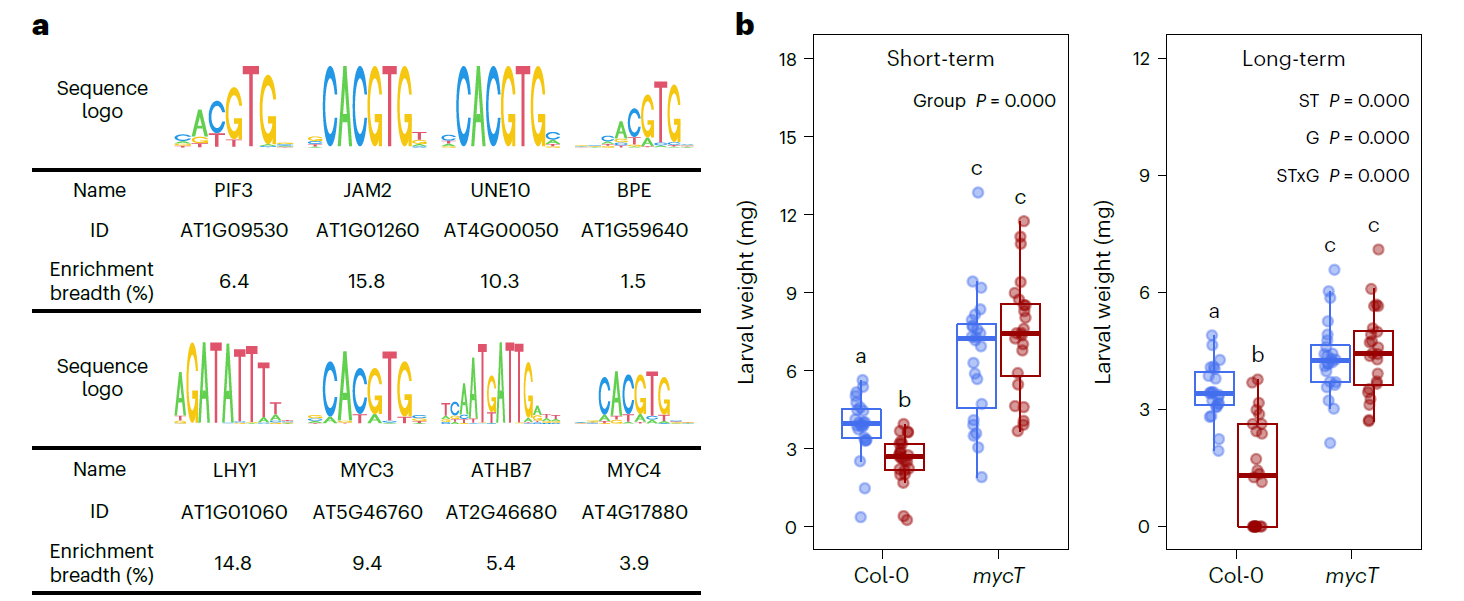
JA诱导的长期抗虫性依赖于转录因子MYC2/3/4
这么多现象,选择哪个进行下一步研究呢?那就选择结果是正向的那个吧。
为了进一步研究JA诱导的长期免疫记忆对棉叶虫Sl保持抗性的转录调控机制,作者选择了203个显著差异表基因,研究其启动子区域(转录起始位点上游1000bp),期望找到与转录因子显著结合的Motif。结果发现显著富集到的Motif大多都含有典型的Motif G-box (CACGTG)。这个Motif的转录因子bHLH的结合位点,包括调控JA的转录因子MYC2/3/4. 定位到关键的转录因子,那么下一步就是验证其功能。于是便利用MYC2/3/4的三重突变体(mycT)与野生型进行比较,分别进行短期的JA诱导和长期的JA诱导。和之前的研究报道一致,哪怕是用水处理三重突变体,棉叶虫的体重与野生型相比均显著增加了,不管是长期诱导还是短期诱导。产生这个结果的一个原因是三重突变体的基础免疫系统被破坏。对野生型植株而言,JA处理后,不管是长期还是短期,棉叶虫Sl的体重均降低了,这说明JA能够诱导野生型植株对棉叶虫Sl的抗性。JA处理对三重突变体均无显著影响,不论是长期还是短期,这表明MYC2/3/4在JA诱导拟南芥植株对棉叶虫Sl的抗性过程中有着至关重要的作用。
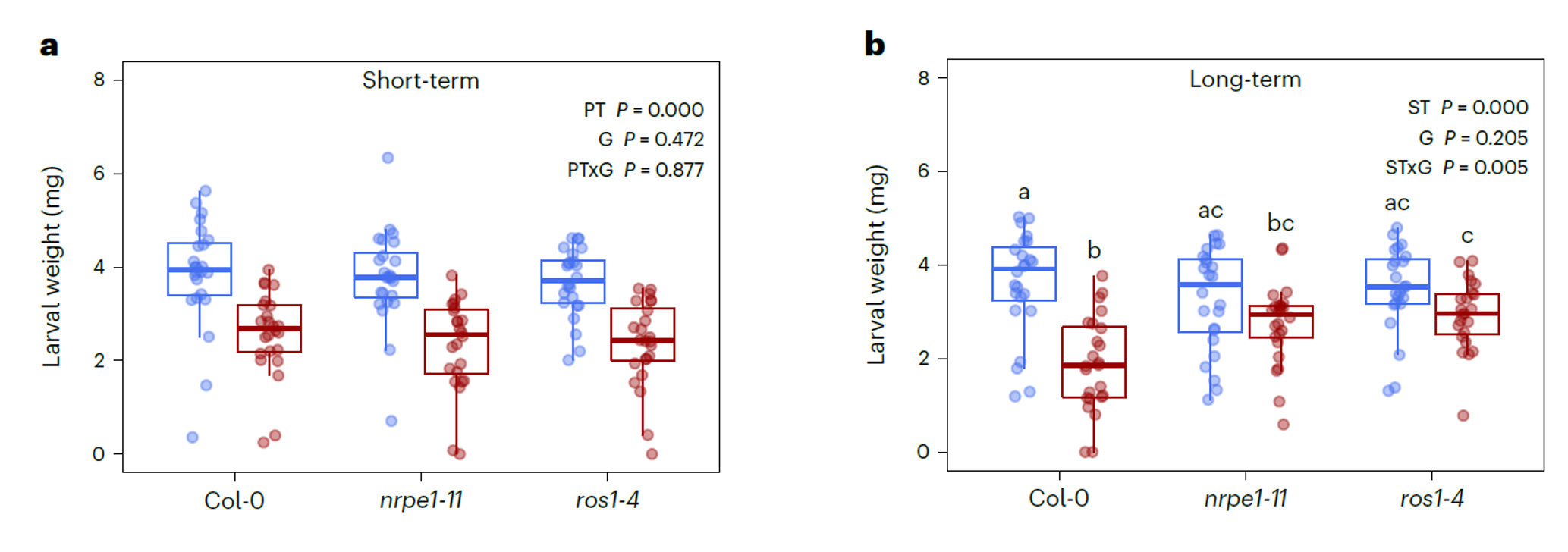
JA诱导的长期抗虫性需要DNA(去)甲基化的参与
幼苗期的拟南芥对棉叶虫Sl是没有抗性的,但是JA处理后3周却产生了抗性,这表明这其中存在一种自我存续的抗性信号,通过细胞分裂的方式传递到新形成的叶片中。在细胞分裂过程中会发生DNA甲基化的变化。先前的研究表明转座元件DNA甲基化的改变能够调控防御相关基因的表达。在拟南芥中,RdDM和DNA去甲基化酶ROS1的拮抗作用共同调控转座元件的DNA甲基化。于是,作者就想探究下这个调节系统是否也参与了JA诱导的短期和长期抗虫性。同样是利用突变体,RdDM突变体是nrpe1-11,ROS1突变体是ros1-4. 短期诱导处理的代码是PT,长期诱导处理的代码是ST。短期诱导的时,突变体和野生型基本是持平的,也就是说棉叶虫Sl不管是吃的野生型植株叶片还是突变体植株的叶片,都没有差异;但是可以看到的是,JA处理与对照相比是有显著的差异的(PT P=0.000)。再看长期诱导,野生型植株在处理前后是有显著差异的,但是两个突变体在处理前后都没有显著的差异了。
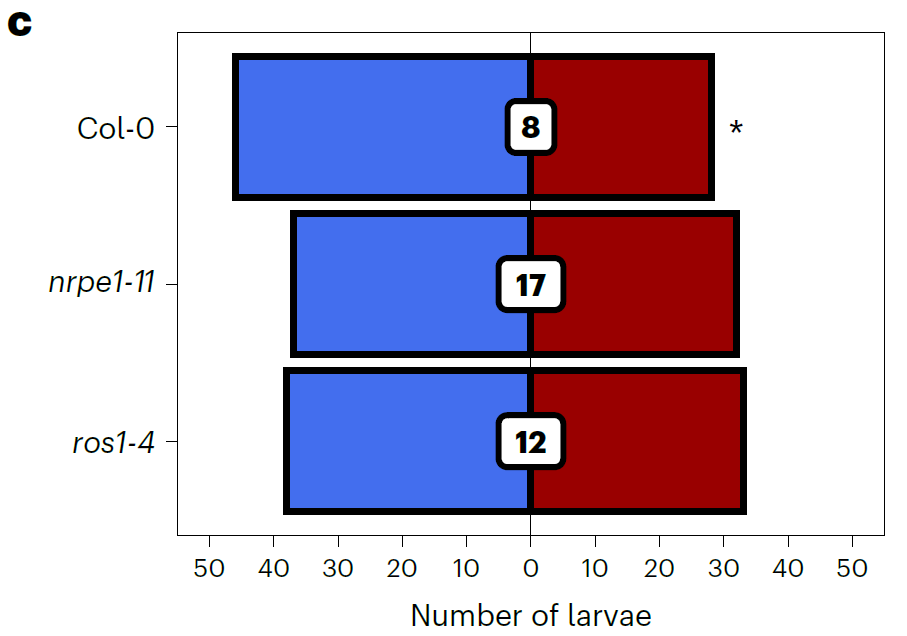
为了进一步探究两个基因突变后对JA长期诱导抗性的影响,作者设计了双向选择试验,也就是将2周六的拟南芥幼苗进行处理,3周后让棉叶虫Sl自由选择吃JA长期诱导后的叶片和对照水处理的叶片。结果发现只有野生型植株在JA长期诱导后具有显著的差异。这个结果和上面的结果共同说明RdDM和ROS1两者很可能都参与了JA诱导的对棉叶虫Sl的长期抗性。
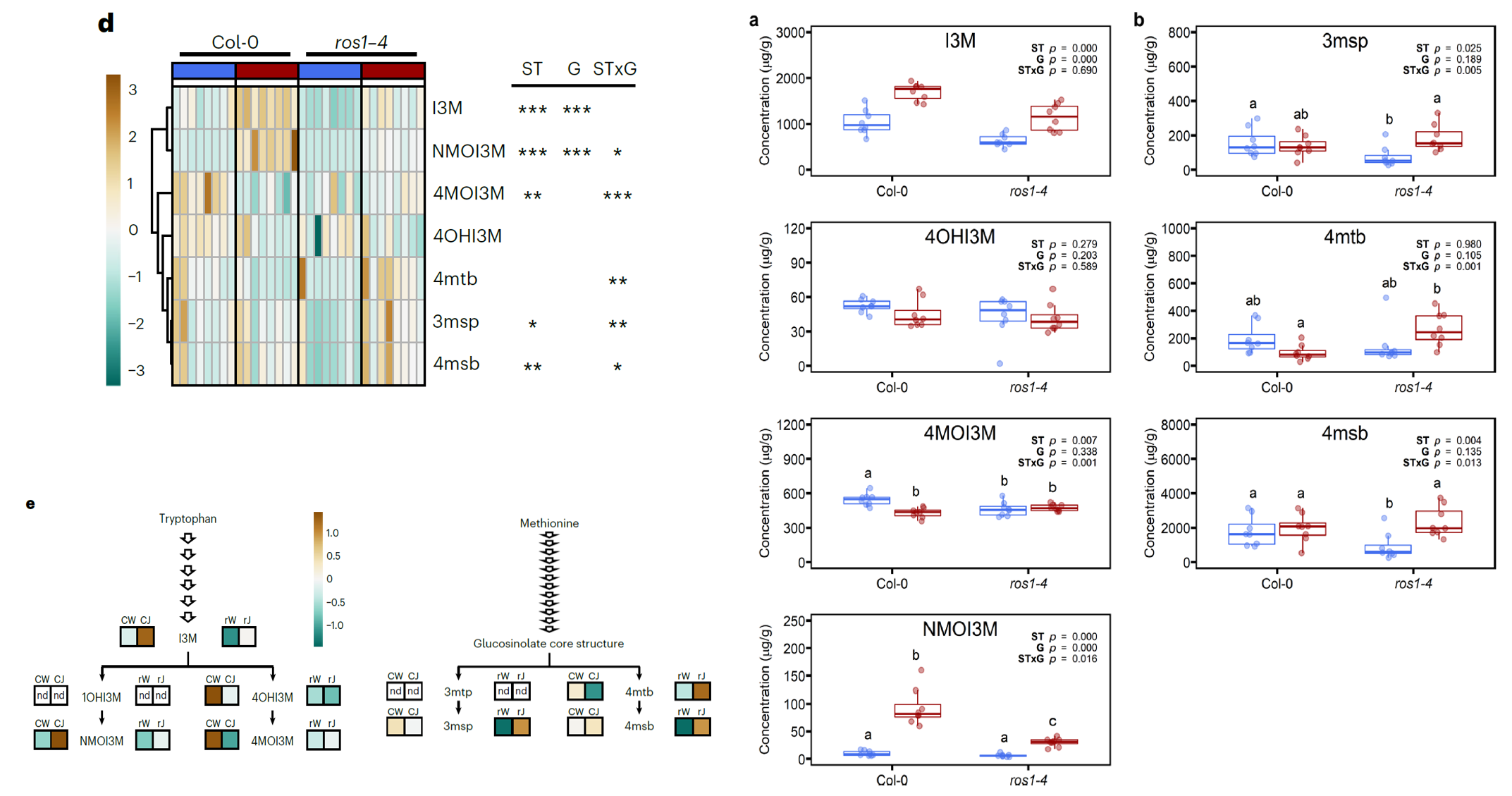
JA诱导的长期免疫与吲哚硫代葡萄糖苷的ROS1依赖性有关
在转录组中鉴定到与JA诱导的对棉叶虫Sl长期抗性相关的203个基因,这203个基因的GO富集分析发现这些基因调控硫代葡萄糖苷的生物合成。再结合上一步发现的DNA甲基化和去甲基化调控JA诱导的长期免疫,作者们就猜想由表观遗传调控的硫代葡萄糖苷池的组成和大小影响了JA诱导的长期免疫。于是呢就用HPLC–QqQ检测JA长期诱导后野生型和突变体ros1-4中硫代葡萄糖苷的含量差异。结果发现JA由奥的长期免疫过程中硫代葡萄糖苷的组成会受到显著影响,尤其是吲哚硫代葡萄糖苷(IGs)的浓度。野生型植株在JA处理后,IGs的主要成分芸苔葡糖硫苷(I3M)及其下游的衍生物新芸苔苷(NMOI3M)的含量显著升高;但是呢IG 4 -甲氧基葡萄糖苷酶(4MOI3M)的含量显著下降了。在突变体ros1-4中,IGs的数量减少,其中I3M和NMOI3M含量降低,4MOI3M甚至都检测不到了。
JA诱导的长期免疫的特征
JA诱导的长期免疫的特征是:转座元件上的发生的可变的低度DANA甲基化。
既然利用突变体发现JA诱导的长期免疫和DNA甲基化相关,那就来个全基因组测序(全基因组亚硫酸氢盐测序,whole-genome bisulfite sequencing,WGBS)。结果发现JA处理后,和对照相比所有的DNA甲基化组分都有降低的趋势,但是没有达到显著水平。从PCA的结果也看不出很明显的差异。

是不是就没有差异,无法往下分析了呢?作者对全局的DNA甲基化进行PCA分析,没有呈现出很好的聚类效果,然后再进行聚类分析。在聚类分析的结果中,全局的DNA甲基是有着明显的聚类差异的,CHG和CHH两种DNA甲基化也是有明显的聚类差异的,CG类型的DNA甲基化没有明显的差异。从这两种分析的结果来看,JA处理后的DNA甲基化与对照相比是有差异的。非CG的DNA甲基化主要发生在转座子序列上,因此作者猜测JA诱导后改变的DNA甲基化主要发生在转座子序列上。
考虑到在PCA结果,JA处理的三个样本间聚类效果较差,作者就用JA处理后的三个样本分别与对照的三个样本进行比较,比如将JA.1与三个对照进行比较。这样分析的话不但可以找到与对照有差异的差异甲基化区域(DMR),还可以找出JA处理的三个重复之间的DMR。分别鉴定到325、291和260个DMR。这些DMR虽然都是比较短的序列,但是呢这些序列都富含胞嘧啶,也展示出DNA甲基化的实质性变化。从整体上来看,这些差异的甲基化区域大多是CHH和CHG,很少是CG类型的。

JA诱导的长期免疫的差异基因的DMR
长期 JA-IR 与差异表达防御基因启动子内的顺式作用 DMR 无关
基因启动子区域的DNA甲基化能够影响转录因子与基因启动子区域Motif的结合。从WGBS的结果可以看出JA处理的三个重复之间存在差异,那么这些DMR会不会集中在启动子区域上的某个共性区域上呢?all-C分组中,发现位于1号染色体的两个consensus DMR,但是呢没有在基因的启动子区域上;将距离从100bp增加到500bp也没有发现新的DMRs。在CG和CHG分组中没有发现consensus DMRs。在CHH分组中,当距离为100bp时间,鉴定到10个consensus DMRs;当距离是500bp时,有25个consensus DMRs被鉴定到。也偶部分consensus DMRs位于基因的启动子区域内,但是呢这些基因都不是转录组中的差异基因。由此,作者们认为RdDM和ROS1在JA诱导产生的长期抗性过程中的作用不是通过调控依赖于MYC2/3/4的防御基因的启动子区域的顺势作用DMRs来实现的。
JA长期处理诱导 ATREP2 转座元件家族特异性的长期低甲基化
最近的研究发现转座元件的DNA低甲基化能够通过反式调控机制影响防御基因的表达。反式激活的机制很多,如源自转座子的siRNA. 但是在JA诱导的长期免疫中,低甲基化的转座元件不太可能产生反式调控的siRNA,因为JA处理后,只有很少的consensus DMRs被鉴定到。同一家族或者相关家族的转座元件具有高度的同源性,作者们就假设同一个分类家族的不同转座元件具有相似的反式激活活性。说干就干,先把数据中在JA处理后显著富集的转座元件(超)家族找出来。果然,找到一个关键的转座元件ATREP2,属于Helitron转座元件家族。这个转座元件在JA处理的三个样本中都被显著富集到。除ATREP2外,JA诱导后,还有一些其他的转座元件家族被富集到(很弱的富集),但是呢没有像ATREP2那样被显著富集到,富集倍数也比较低,在三个重复之间也没有一致性。
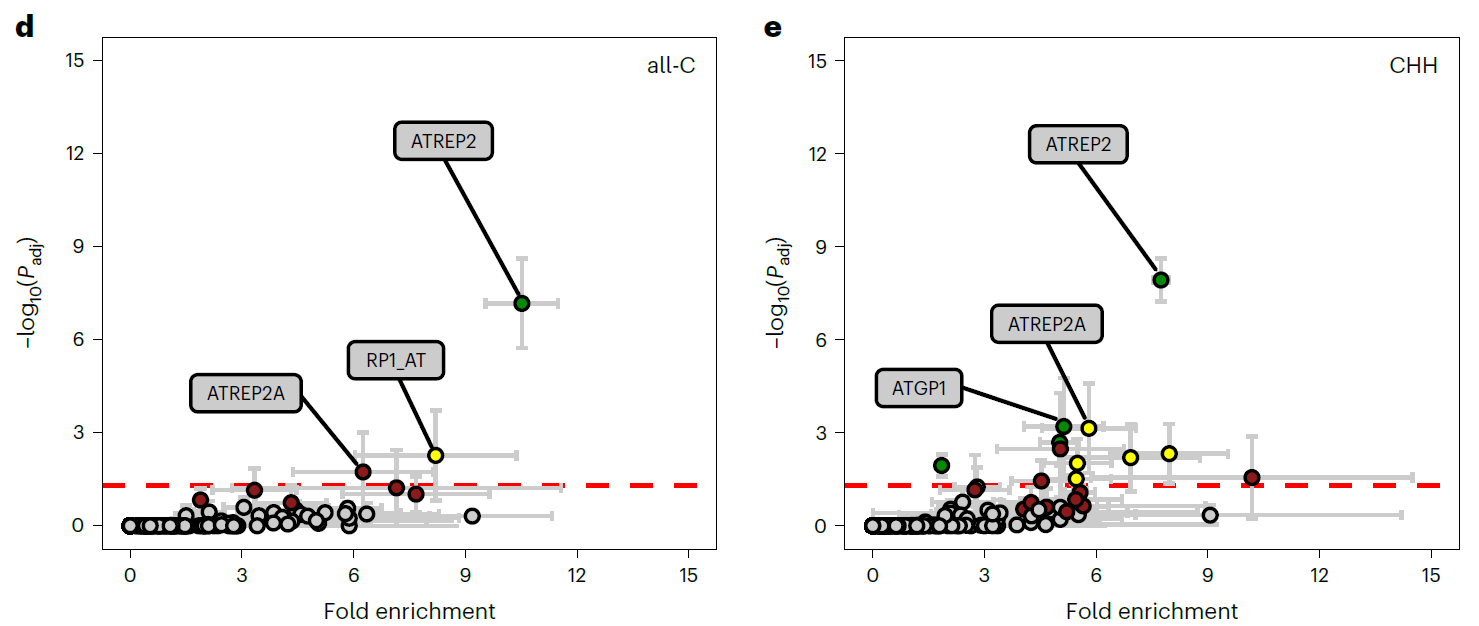
进一步发现ATREP2 DMRs中的大多数都发生了低甲基化,并且分布在所有染色体上。但是在差异表达基因附近,没有发现ATREP2 DMRs。
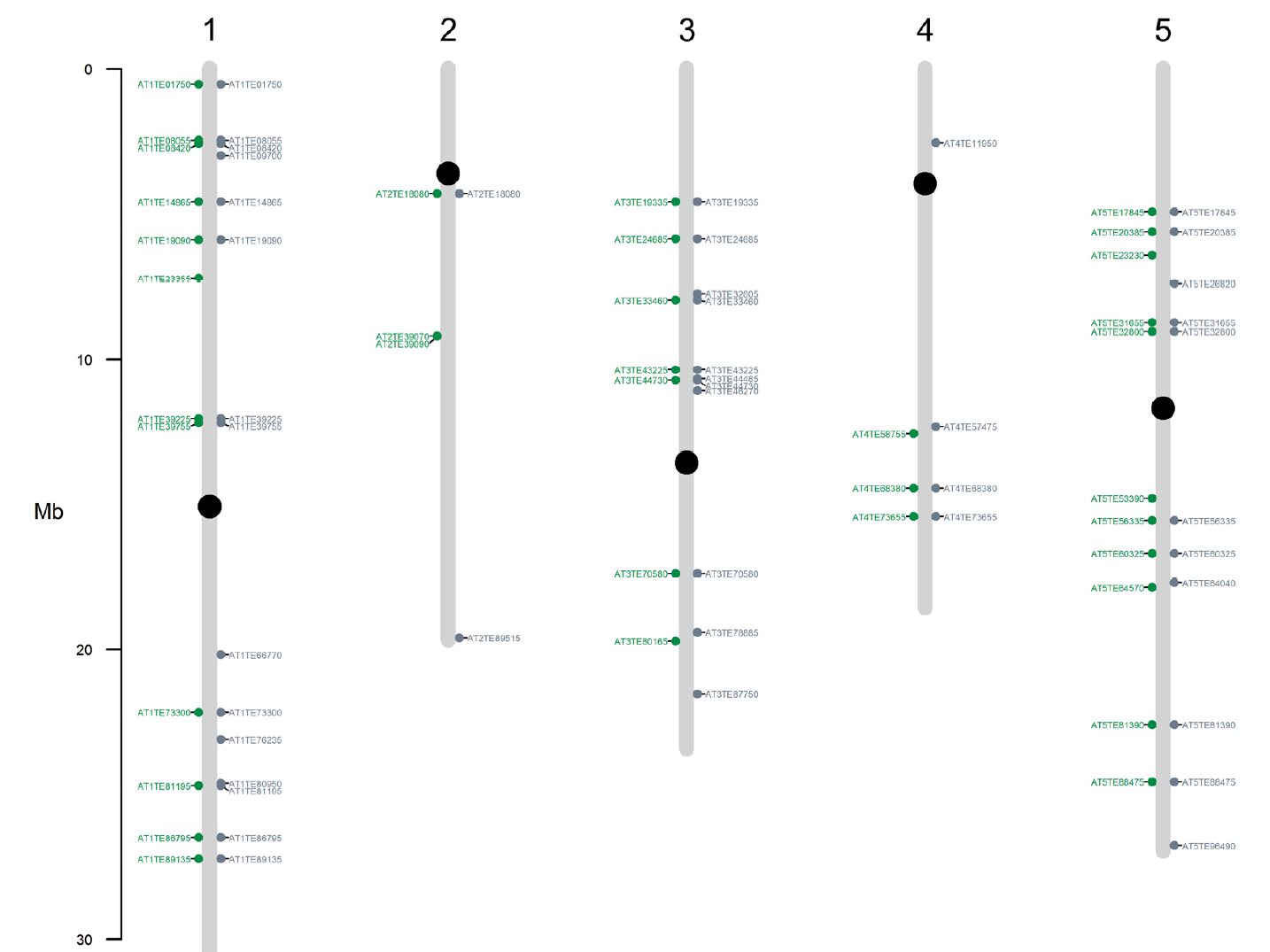
在CG和CHG分组中,JA诱导后富集到的DMRs是LTR反转录转座子的Gypsy超家族。但是呢处于显著富集阈值附近,也么有翻译成特定的TE家族(不是很理解这句话)。
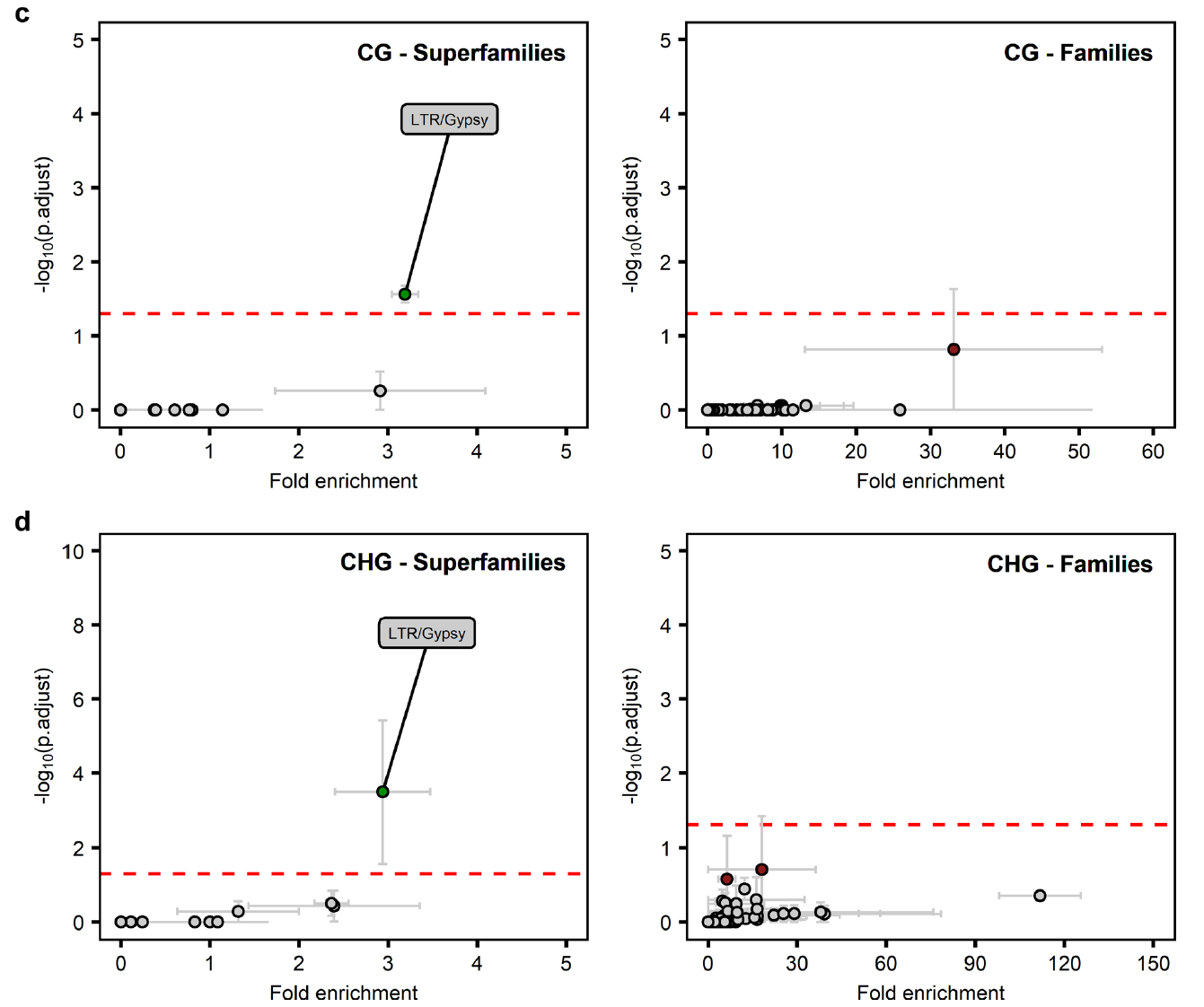
因此,尽管DNA低甲基化存在着变异(差异),但是呢JA诱导的DNA甲基化始终靶向ATREP2家族的转座元件。 结合前面的MYC2/3/4被诱导表达,以及RdDM和ROS1在甲基化中的核心功能,作者认为他们的WGBS结果表明ATREP2 TE家族成员的随机低甲基化通过反式作用机制诱导和/或启动JA依赖的防御基因的表达。
RdDM调控ATREP2转座子的DNA甲基化
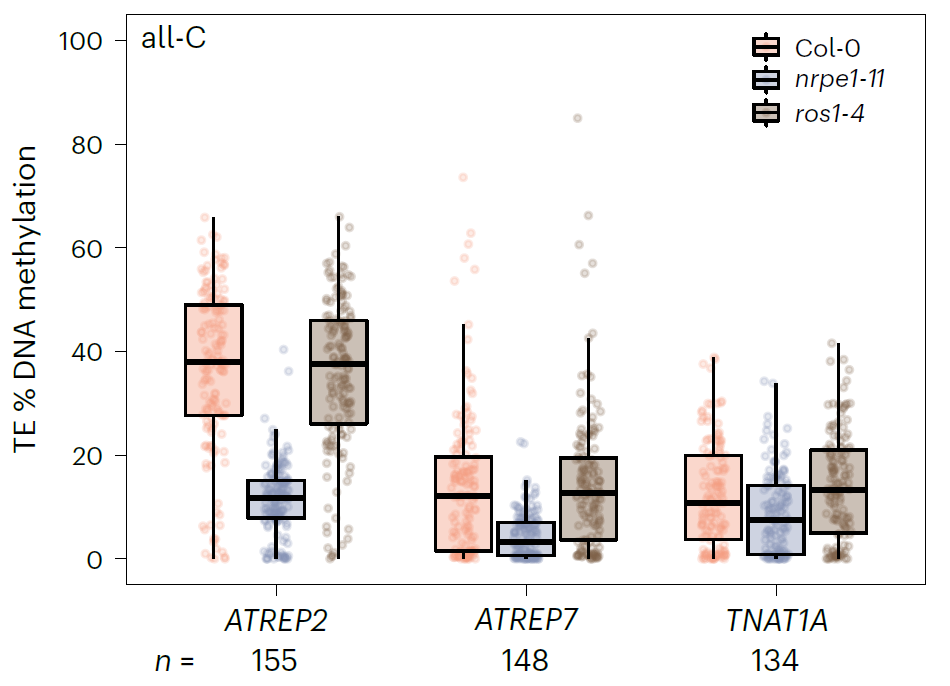
既然都定位到了ATREP2转座元件家族受DNA甲基化的影响,那么下一步就是探究其中的调控机制。先前的研究根据依赖ROS1的去甲基化的拮抗程度对拟南芥RdDM靶位点的进行了分类。通过比较分析这些数据发现164个ATREP2转座元件家族成员中的25个和之前鉴定的RdDM位点重合。但是呢这个研究中鉴定到的受JA诱导调控的大部分DMRs与RdDM的位点并不吻合。先前的研究结果发现JA诱导的长期抗性依赖于RdDM和ROS1,而且JA处理对CHH类型的甲基化有着很明显的影响。现在却发现受JA诱导处理的DMRs却和RdDM的位点无法对应上,这是令人感到惊讶的。这个时候怎么办呢?直接对突变体进行甲基化测序,分别测了野生型、RdDM突变体nrpe1-11和ROS1突变体ros1-4. 选择三个大小差不多的转座子家族进行比较,即具有164个成员的ATREP7和162个成员的TNAT1A,这两个转座元件家族都和JA处理后的DNA甲基化无关;第三个转座元件家族就是ATREP2.结果发现ATREP2在野生型植株中被高度甲基化。而且,在突变体nrpe1-11中,ATREP2转座元件的甲基化水平急剧降低;但是在突变体ros1-4中甲基化水平还是很高。这表明JA处理野生型植株后,ATREP2富集的DMRs表现出高度依赖于RdDM的DAN甲基化。
ROS1靶向ATREP2进行DNA去甲基化
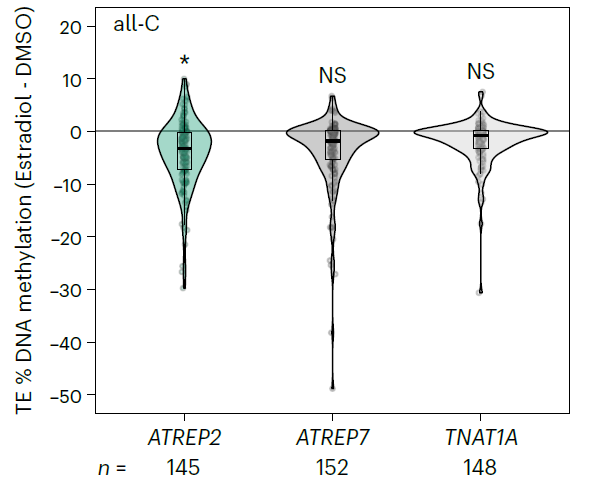
为什么在上面这个图里面突变体ros1-4的甲基化水平和野生型植株的差不多呢?也就为什么突变体ros1-4没有对ATREP2的甲基化水平产生影响呢?在突变体nrpe1-11中ATREP2的甲基化水平降低了,既然RdDM和ROS1具有拮抗作用,那么团变体ros1-4中ATREP2的甲基化水平也应该有很明显的变化啊。为什么会这样呢?作者猜测是由于ATREP2的甲基化水平在野生型植株中已经达到饱和了。为了验证ROS1是否能够靶向ATREP2转座子进行DNA甲基化,首先是利用β -雌二醇诱导的YFP标记ROS1,然后利用三代测序检测ROS1对ATREP2甲基化的影响水平。值得注意的是,与来自两个对照家族的TEs相比,雌二醇在ATREP2转座元件中诱导了广泛的DNA低甲基化。异位诱导后的ROS1靶向ATREP2使得DAN去甲基化再次证明ATREP2的甲基化是受Rd和ROS1的拮抗调控的。
AGO1在JA诱导长期免疫中的功能
来自ATREP2转座子的sRNA与AGO1关联,这个AGO1呢在JA诱导的长期抗虫性中具有重要作用。
最近的研究发现AGO1能够与SRNAs关联并通过改变基因的染色质结构和募集Pol II来诱导依赖于JA的免疫基因的表达。
为了验证来自于ATREP2转座子的与AGO1关联的sRNA在JA诱导的长期免疫中是否发挥功能,作者们分析了来自茉莉酸甲酯处理后的来自的AGO1核的sRNA测序数据,结果发现来自于ATREP2的sRNA被显著富集到,尤其是长度为21nt的RNA。这些来自于ATREP2的sRNA中,$\frac{21nt RNA}{24nt RNA}$的比例明显升高,说明茉莉酸甲酯降低了Pol IV依赖的沉默,增加了Pol II转录的ATREP2 TEs产生PTGS依赖的21nt sRNA。但是呢这些21nt RNA和24nt RNA的表达量均比对照的低,这就和作者们猜想的JA诱导提来自于ATREP2转座子的21nt sRNA活性不一样。但是呢,值得注意的是,这个树数据来源的研究中的处理和作者们这个文章的处理差很远,数据来源的研究中只用的拟南芥幼苗是10天大的,而且处理1天后就采样了。
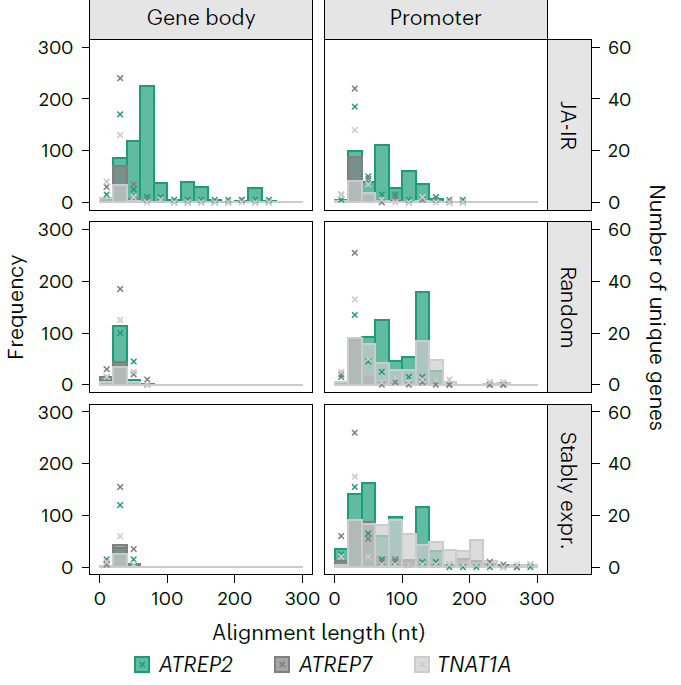
为了找到更多的证据证明来自于ATREP2的sRNA能够反式调控依赖于JA的基因表达, 作者们利用BLAST分析JA长期诱导后显著变化的203个基因和ATREP2的同源性。结果发现,这种富集的序列互补性在IR相关基因的基因体中尤为明显,在随机选取的203个对照基因和203个稳定表达的基因中缺失。
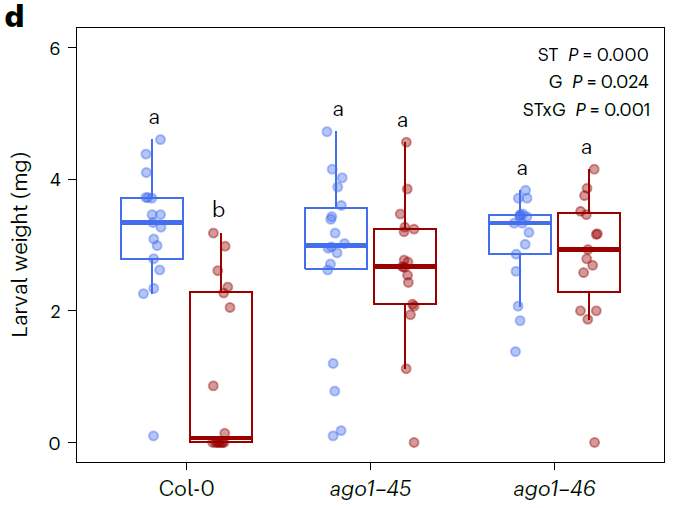
在最后,作者们还测试了AGO1在JA诱导从长期抗虫性中的功能。与野生型植株相比,两个突变体植株都没有展现出显著的JA诱导的长期抗虫性。因此,JA诱导的长期抗虫性需要完整的AGO1蛋白的参与。
研究结论
来自低甲基化的ATREP2的sRNA能够与AGO1结合,对启动或诱导近端/远端依赖于JA的防御基因的表达,并介导JA诱导的长期抗虫性。