氨基酸转运子调控植物免疫-生长平衡
背景知识
先天免疫与诱导免疫
植物的先天免疫可以分为PTI和ETI。除了先天免疫之外,当植物从生物胁迫中恢复以后,会获得持续时间较长的抗性,这种叫做诱导抗性(induced resistance,IR)。获得IR的植株在遭受第二次侵染时,能够更快更强地做出响应以保护自己。IR通常有如下几种方式:
- 由先天免疫激发;
- 由根际微生物激发:如有益微生物;
- 由特定化合物激发:如挥发性有机化合物和β 氨基酸。
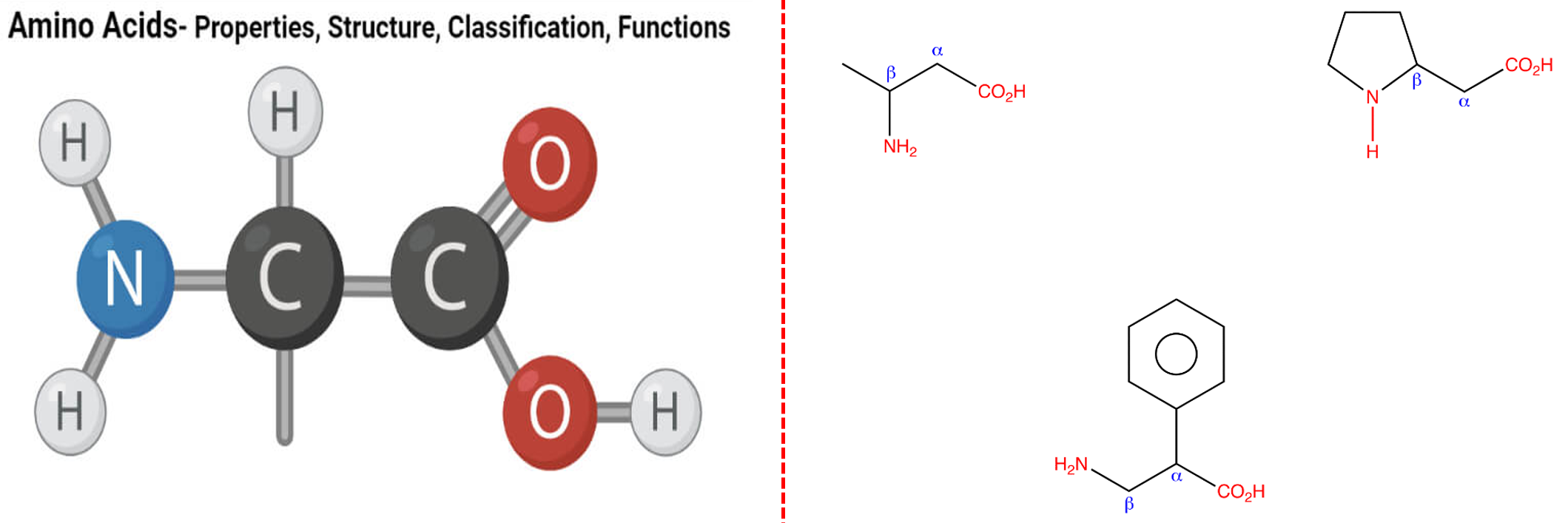
$\beta$氨基酸
我们常见的氨基酸的氨基是在$\alpha$位上的,如果氨基在$\beta$位上就是$\beta$氨基酸。
BABA与RBH

β 氨基丁酸(BABA)激发植物的防御机制可以通过SA途径实现,也可以不需要SA途径的参与。植物收到生物/非生物胁迫时会有大量的β 氨基丁酸积累。先前的研究发现BABA的受体是一个天冬氨酰 tRNA 天冬氨酸酶(INI1)。BABA也是根际信号,刺激植物获得诱导系统抗性。但是胞内转运机制还不清楚。虽然没BABA具有诱导抗性的功能,但是高浓度的BABA会抑制植株的生长。为了找到和BABA同样具有诱导功能的类似物,作者们筛选到了R-β-高丝氨酸(RBH)。RBH能够不依赖于IBI1酶实现对植株的抗性的诱导,而且对植株的生长没有影响。当拟南芥受到Hyaloperonospora arabidopsidis(Hpa,霜霉菌)侵染时,RBH不会激发依赖于SA的基因表达,但是会刺激camalexin的合成;当拟南芥受到Plectosphaerella cucumerina侵染时,RBH会激发依赖于JA途径的防御基因的 表达。由此可见,RBH诱导植物产生抗性的途径和BABA不一样。而且,RBH是被感知和吸收的分子机制还不清楚。
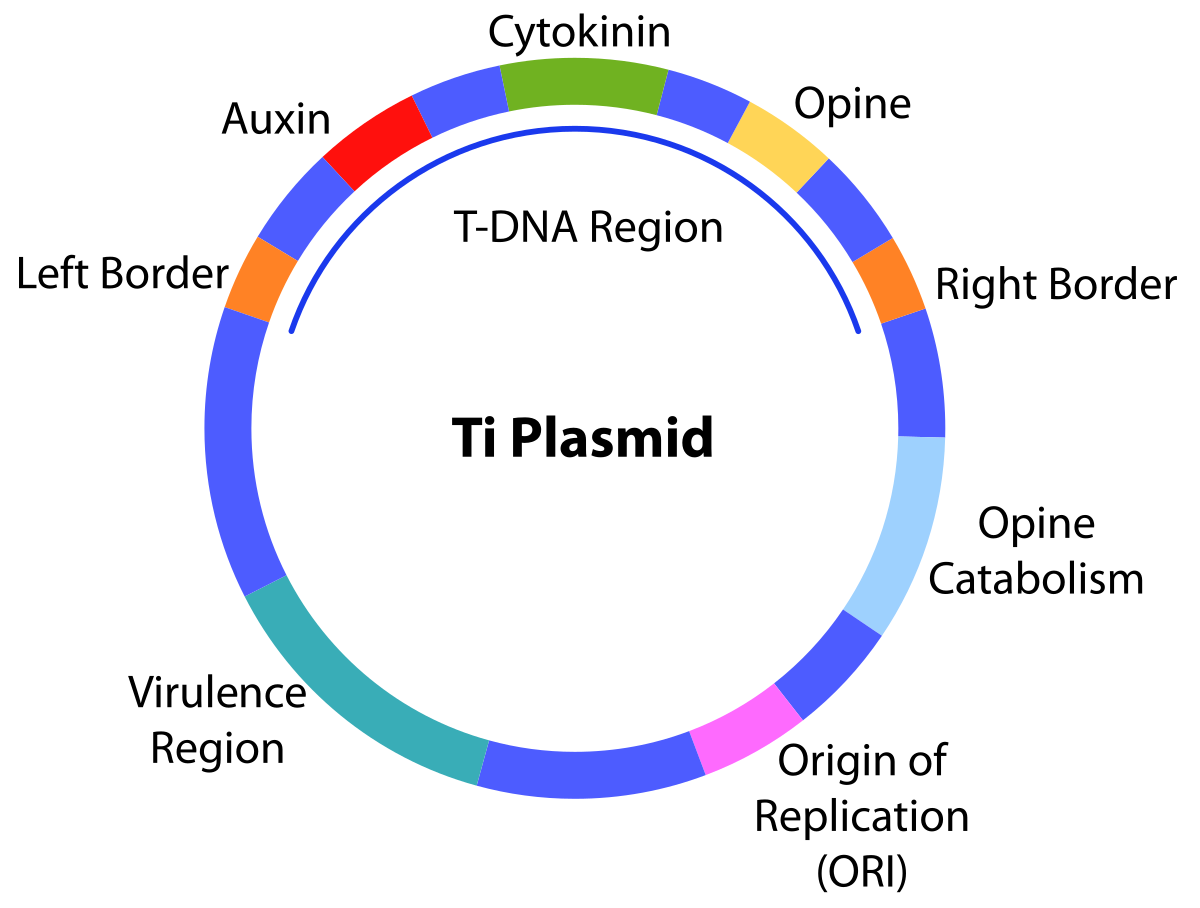
T-DNA
Ti 质粒是土壤农杆菌的天然质粒,该质粒上有一段特殊的 DNA区段,当农杆菌侵染植物细胞时,该 DNA 区段能自发转移,插入植物染色体 DNA中,Ti 质粒上的这一段能转移的DNA被叫做 T-DNA。人们根据这一现象,将Ti质粒进行改造,将感兴趣的基因放进T-DNA 区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化。
结果分析
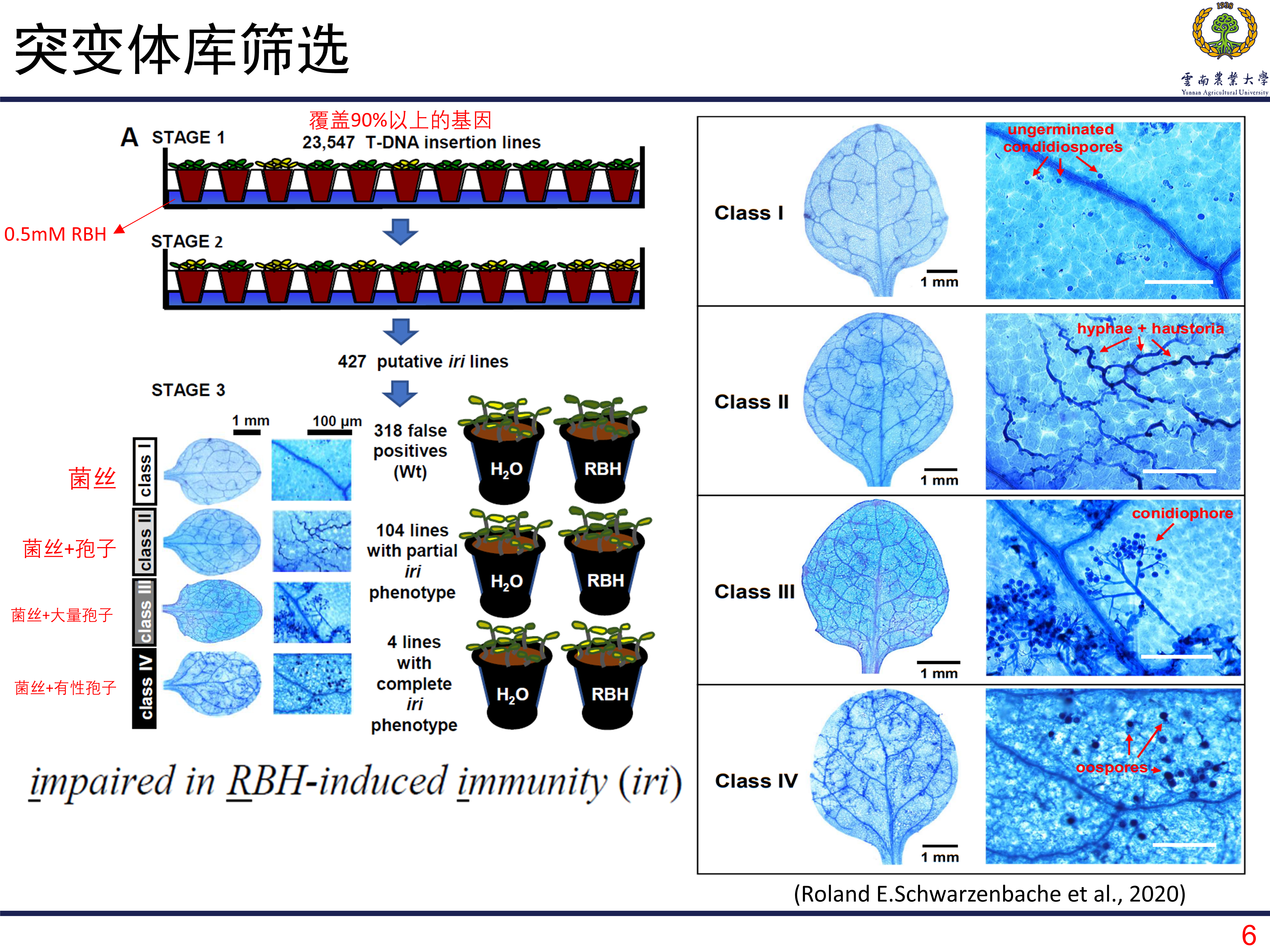
突变体库筛选靶标基因
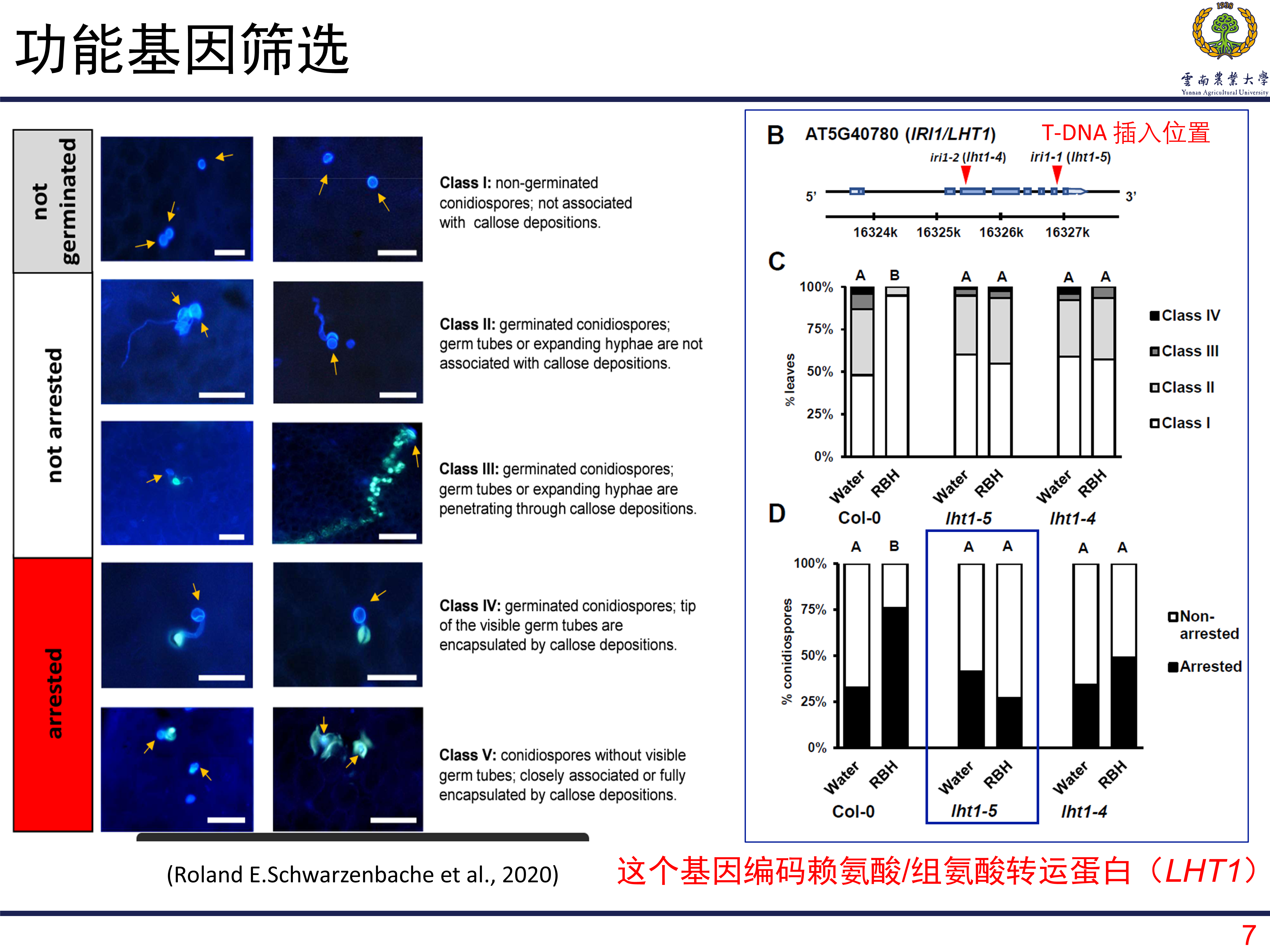
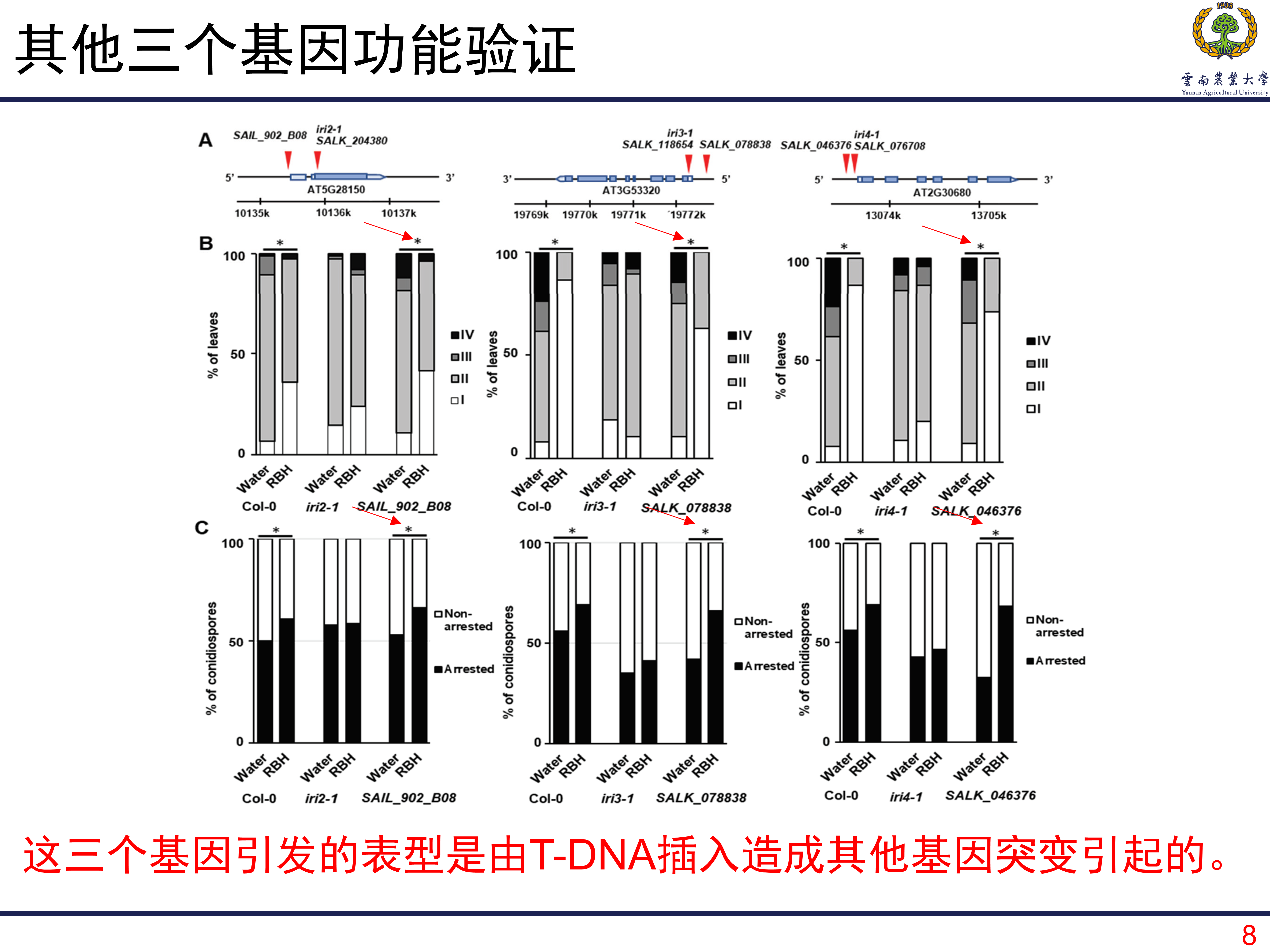
如何定位和和RHB诱导的抗性相关的基因呢?T-DNA筛选是一种很有效的方法,得益于前期的研究,作者从一个特殊的T-DNA插入系中筛选目的基因。一共是有23547个突变系。将这些突变系种植在土壤中,土壤用浓度为0.5mM的RBH进行浸泡至饱和,然后接种病原菌Hpa。如果某个株系在接种后表现出明显的发病,那么对应的株系就很有可能是目的基因所在的株系。经过两轮筛选后,一共筛选到427个候选的株系。在此基础上,利用台盼蓝染色观察处理和对照之间的病原菌生长情况。其中有104个展现出部分功能缺失,有四个株系表现为完全的功能缺失,就选择这四个株系所包含的关键基因作为目的基因进行研究。
在对第一个基因进行研究的时候发现,这个基因在敲除系中表现出更差的抗性。但是在对其他三个基因进行研究的时候没有发现这现象,甚至在敲除后植株的抗性更高了。那么也就将第一个基因确定为目标基因,研究发现这个基因编码赖氨酸/组氨酸转运子,名为为LHT1。
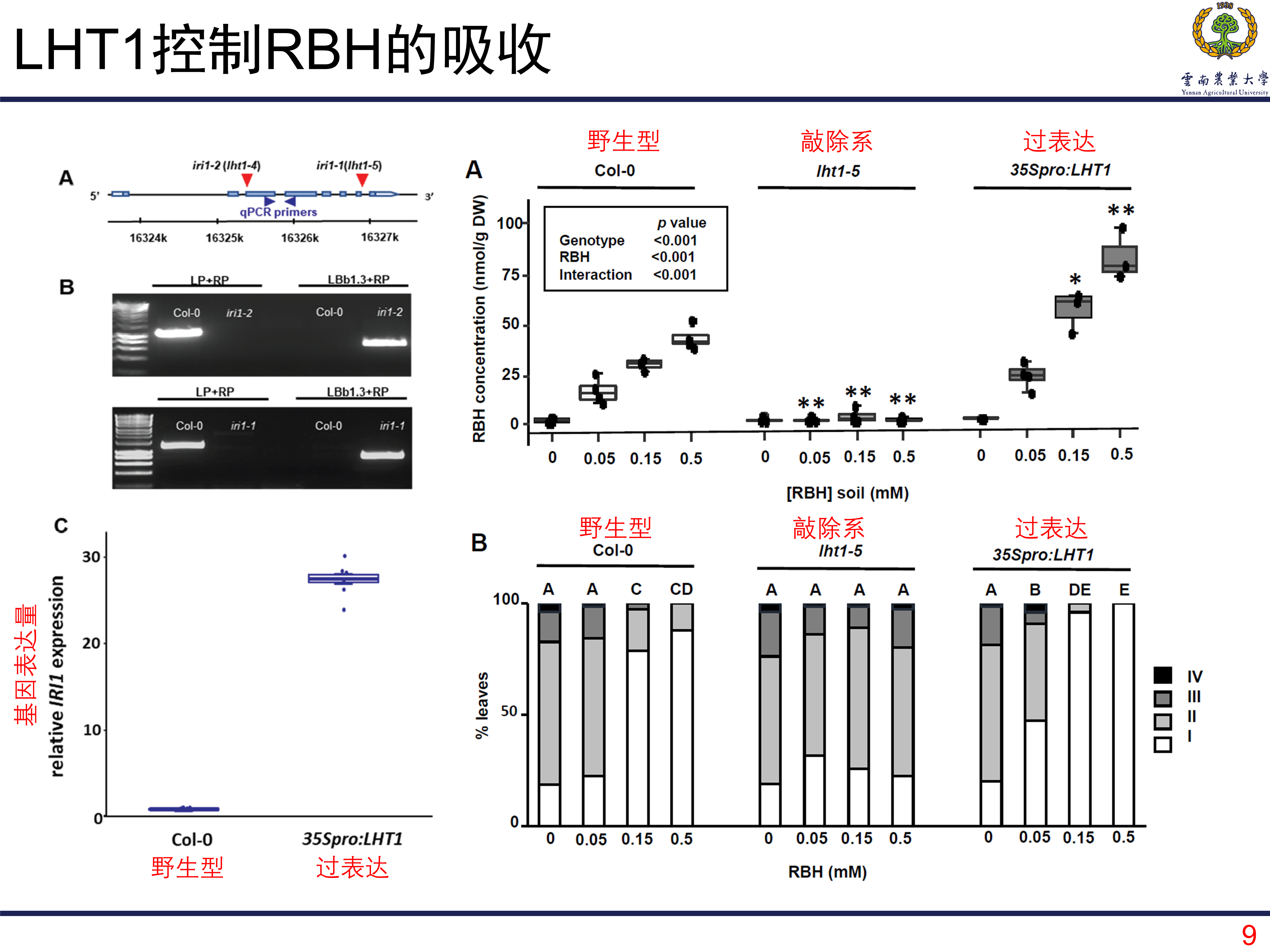
LHT1调控植株吸收RBH
首先我们知道的是,RBH能够诱导植株的免疫,那LHT1是如何调控RBH的呢?LHT1能够转运酸性及中性氨基酸,那首先想到的是LHT1是不是通过调控植株对RBH的吸收来实现诱导抗性的呢?为了探究这个问题,那就是典型的敲除和过表达。首先可以看到的是,敲除系对RBH的吸收明显受到抑制;而过表达系能够吸收更多的RBH,比野生型还多。这个结果说明LHT1调控植株对RBH的吸收。而且对应的接种结果也一样。
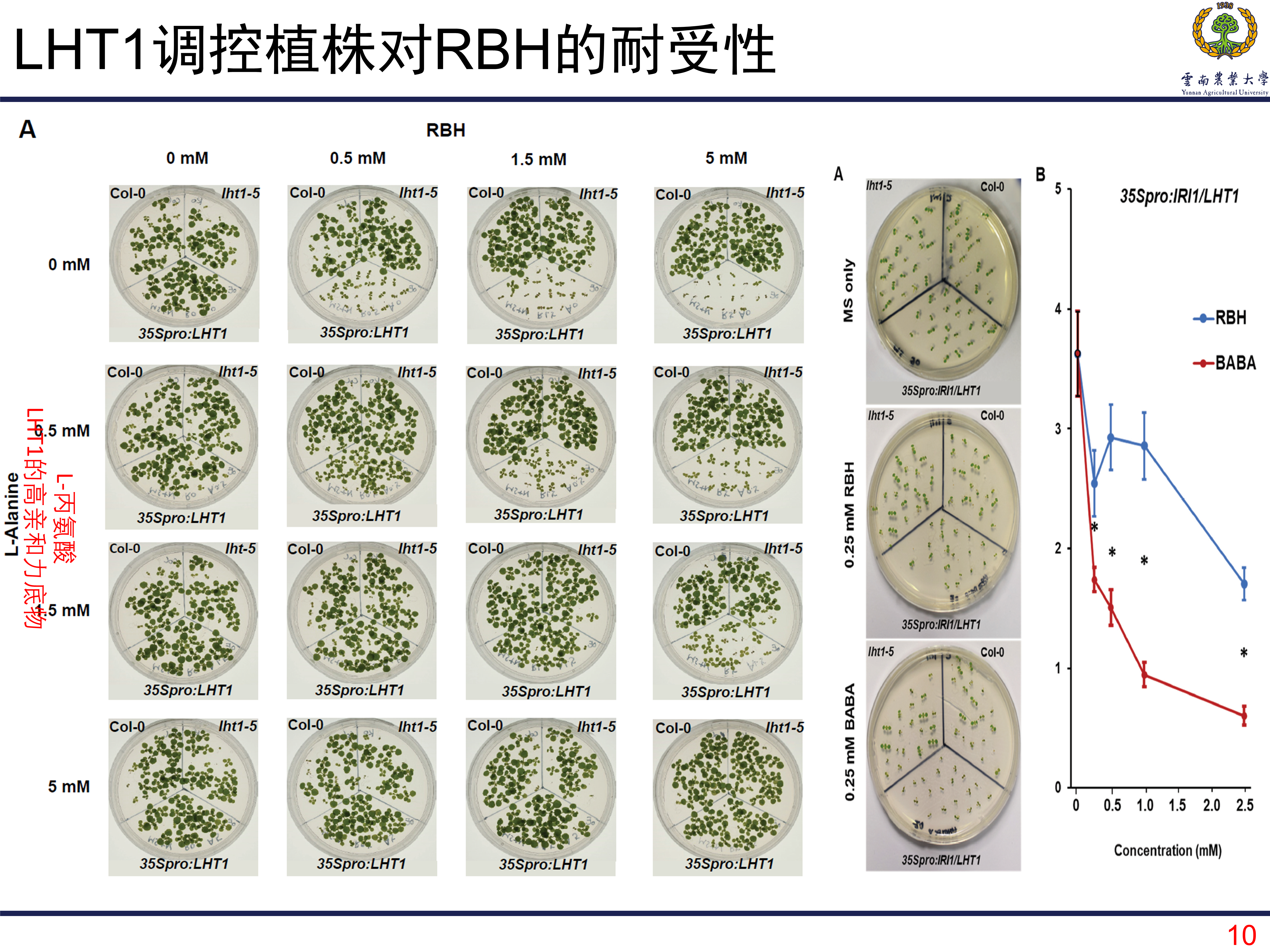
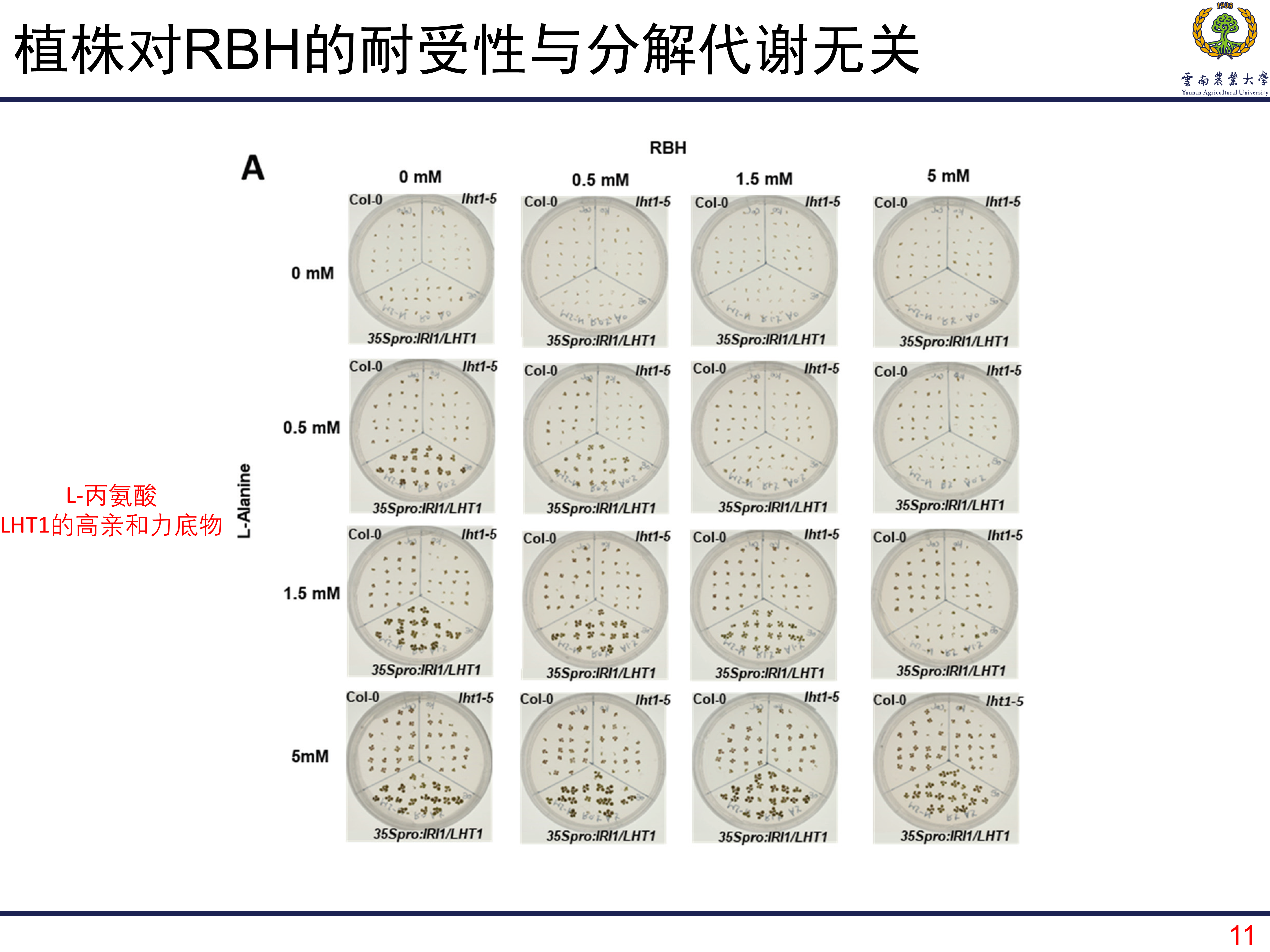
虽然RBH不像BABA那样明显影响植株的生长,但是对植株的生长还是有那么一些影响的。那么植株对RBH的耐受性是不是由LHT1调控的呢?首先可以看到的是,当RBH的浓度越来越高以后,植株的生长受到了明显的抑制。但是,当往培养基中加入LHT1的高亲和力底物L-丙氨酸以后,这种抑制作用得到明显的缓解。这个结果表明,植株对RBH的耐受性是由LHT1控制的,当LHT1吸收更多的RBH,那么植株受到的毒害就越严重。总之,是LHT1调控植株吸收RBH,当RBH浓度过高时,生长就会受到明显的抑制。
RBH分解代谢与耐受性
有没有这样一种可能?拟南芥植株能够更好地将RBH进行分解代谢,这样呢RBH的毒害性就降低了。为了验证这种可能性,作者利用无N培养基,让植株吸收的RBH成为唯一的N源,如果植株能够正常代谢RBH,那RBH就会被代谢成N源,植株也就能够正常生长。但是,研究发现拟南芥并不能代谢RBH。但是当培养基中由L-丙氨酸的时候,植株是可以正常生长的,而且过表达的植株长得更好。这个结果说明植株对RBH的耐受性并不是因为植株能够分解代谢RBH,而是植株通过表达LHT1控制对RBH的吸收。
LHT1调控植株对BABA的吸收
BABA和RBH在结构上很像,那LHT1能不能调控植株对BABA的吸收呢?类似的实验证明是可以的,完全可以。而且过表达植株在0.025mM的浓度的时候就有很好的诱导效应,浓度再增加对诱导免疫没有多少改善。
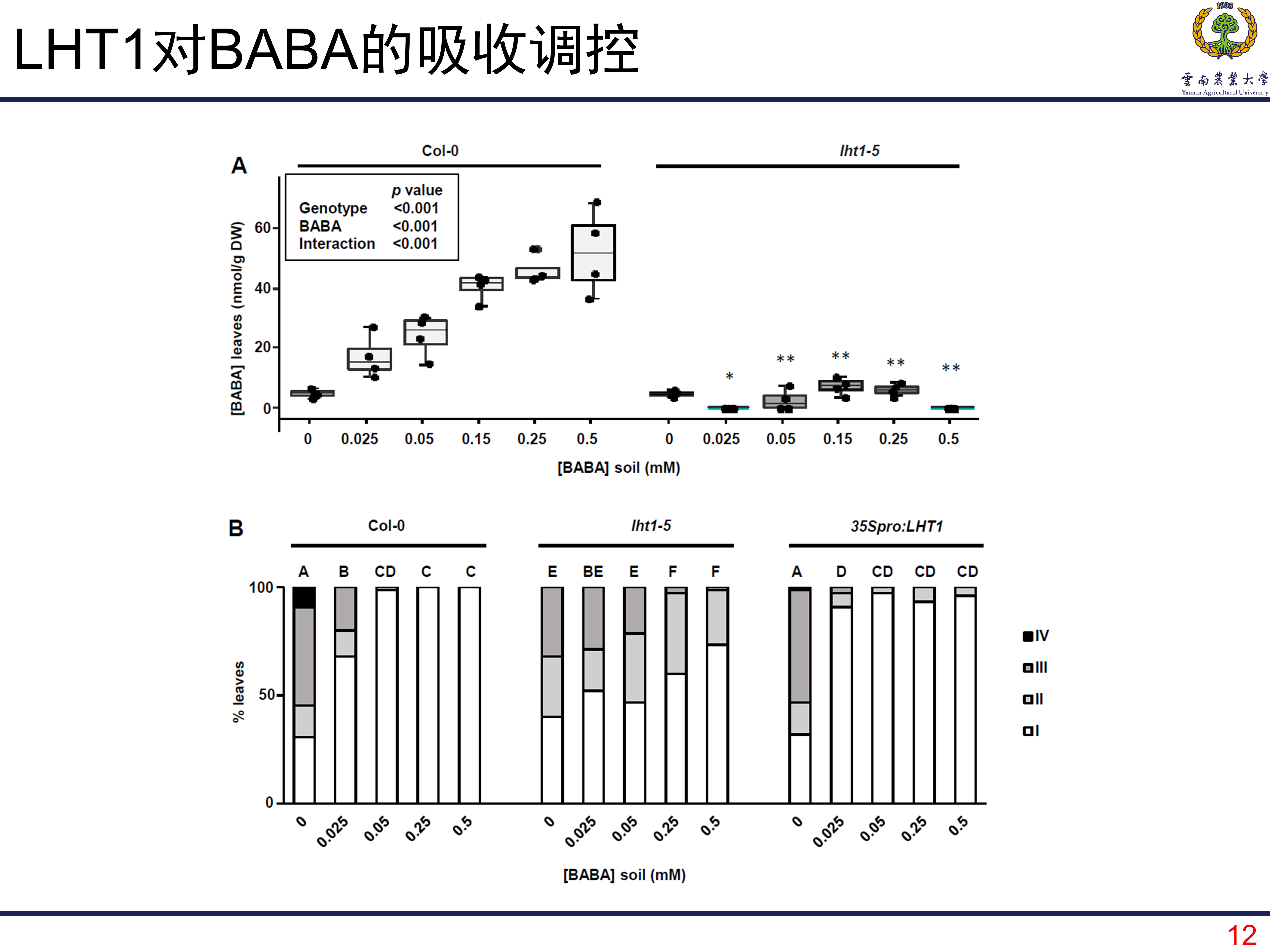
LHT1调控植株对BABA耐受性
和先前的验证方法一样,验证LHT1能否影响植株对BABA的耐受性。结果和预料的一样,BABA浓度升高后,毒性越明显;过表达的植株所受到的毒害效应更明显。

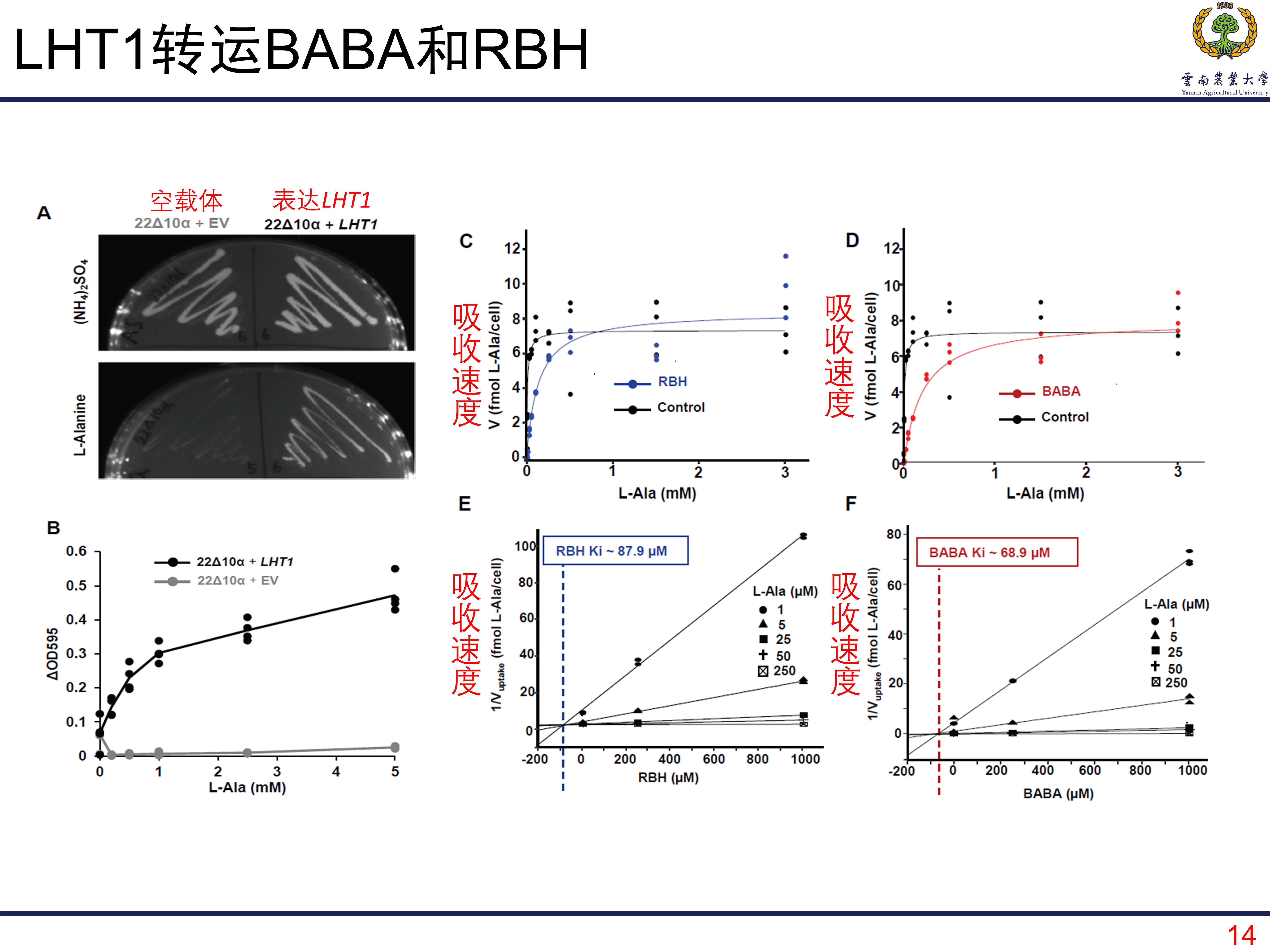
LHT1对BABA和RBH的转运
首先是利用一个特殊的酵母再次证明了LHT1能够调控氨基酸的吸收。在此基础上呢,像测试LHT1对BABA和RBH的转运动力学(简单来说就是看看浓度到那个地方的时候转运的量就没有明显的变化了)。从下图的C和D可以看到的是,加入BABA以后,转运的效率和总量并没有很明显的变化;但是,当加入RBH以后,L-丙氨酸低浓度的时候,表面有增强转运,随着L-丙氨酸浓度升高,转运效率得到明显的提高,而且平台期也来得更晚一些。这也就是表明RBH能够促进L-丙氨酸的吸收。
阅读收获